Region of Terminal Digestion and Absorption: The Intestine
Region of Terminal
Digestion and Absorption:
The Intestine
The importance of the intestine varies widely among animal groups. In invertebrates that have extensive digestive diverticula in which food is broken down and phagocytized, the intestine may serve only as a pathway for conducting wastes out of the body. In other invertebrates with simple stomachs, and in all vertebrates, the intestine is equipped for both digestion and absorption.
Devices for increasing the internal surface area of the intestine are highly developed in vertebrates, but are generally absent among invertebrates. Perhaps the most direct way to increase the absorptive surface of the gut is to increase its length. Coiling of the intestine is common among all vertebrate groups and reaches its highest development in mammals, in which the length of the intestine may exceed eight times the length of the body. Although a coiled intestine is rare among invertebrates, other strategies for increasing surface sometimes occur. For example, the typhlosole of terrestrial oligochaete worms (see Figure 17-12), an inward folding of the dorsal intestinal wall that runs the full length of the intestine, effectively increases internal surface area of the gut in a narrow body lacking space for a coiled intestine.
Lampreys and sharks have longitudinal or spiral folds in their intestine. Other vertebrates have developed elaborate folds (amphibians and reptiles) and minute fingerlike projections called villi (birds and mammals), which give the inner surface of fresh intestinal tissue the appearance of velvet (Figure 34-12). The electron microscope reveals that each cell lining the intestinal cavity additionally is bordered by hundreds of short, delicate processes called microvilli (Figure 34-13C and D). These processes, together with larger villi and intestinal folds, may increase the internal surface area of the intestine more than a million times as compared to a smooth cylinder of the same diameter. This elaborate surface greatly facilitates the absorption of food molecules.
Digestion in the Vertebrate Small Intestine
Food is released into the small intestine through the pyloric sphincter, which relaxes at intervals to allow entry of acidic stomach contents into the initial segment of the small intestine, the duodenum. Two secretions pour into this region: pancreatic juice and bile (Figure 34-14). Both of these secretions have a high bicarbonate content, especially pancreatic juice, which effectively neutralizes gastric acid, raising the pH of the liquefied food mass, now called chyme, from 1.5 to 7 as it enters the duodenum. This change in pH is essential because all intestinal enzymes are effective only in a neutral or slightly alkaline medium.
Cells of the intestinal mucosa, like those of the stomach mucosa, are subjected to considerable wear and are constantly undergoing replacement. Cells deep in the crypt between adjacent villi divide rapidly and migrate up the villus. In mammals the cells reach the tip of the villus in about twodays. There they are shed, along with their membrane enzymes, into the lumen at the rate of some 17 billion a day along the length of the human intestine. Before they are shed, however, these cells differentiate into absorptive cells that transport nutrients into the network of blood and lymph vessels, once digestion is complete.
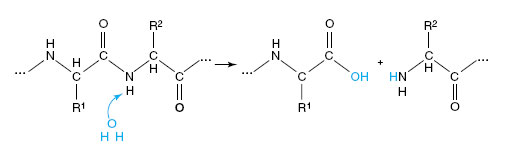
Pancreatic Enzymes The pancreatic secretion of vertebrates contains several enzymes of major importance in digestion (Figure 34-14). Two powerful proteases, trypsin and chymotrypsin, continue enzymatic digestion of proteins begun by pepsin, which is now inactivated by the alkalinity of the intestine. Trypsin and chymotrypsin, like pepsin, are highly specific proteases that split apart peptide bonds deep inside the protein molecule. The hydrolysis of a peptide linkage may be shown as: Pancreatic juice also contains carboxypeptidase, which removes amino acids from the carboxyl ends of polypeptides; pancreatic lipase, which hydrolyzes fats into fatty acids and glycerol; pancreatic amylase, a starch-splitting enzyme identical to salivary amylase in its action; and nucleases, which degrade RNA and DNA to nucleotides.
Membrane Enzymes The cells lining the intestine have digestive enzymes embedded in their surface membrane that continue digestion of carbohydrates, proteins, and phosphate compounds (Figure 34-14). These enzymes of the microvillus membrane (Figure 34-13D) include aminopeptidase that splits terminal amino acids from the amino end of short peptides, and several disaccharidases, enzymes that split 12-carbon sugar molecules into 6-carbon units. The disaccharidases include maltase, which splits maltose into two molecules of glucose; sucrase, which splits sucrose to fructose and glucose; and lactase, which breaks lactose (milk sugar) into glucose and galactose. Also present is alkaline phosphatase, an enzyme that attacks a variety of phosphate compounds.
Although milk is the universal food of newborn mammals and one of the most complete human foods,many adult humans cannot digest milk because they are deficient in lactase, the enzyme that hydrolyzes lactose (milk sugar). Lactose intolerance is genetically determined. It is characterized by abdominal bloating, cramps, flatulence, and watery diarrhea, all appearing within 30 to 90 minutes after ingesting milk or its unfermented by-products. (Fermented dairy products, such as yogurt and cheese, create no intolerance problems.)
Northern Europeans and their descendants, which include the majority of North American whites, are most tolerant of milk. Many other ethnic groups are generally intolerant to lactose, including the Japanese, Chinese, Jews in Israel, Eskimos, South American Indians, and most African blacks. Only about 30% of North American blacks are tolerant; those who are tolerant are mostly descendants of slaves brought from east and central Africa where dairying is traditional and tolerance to lactose is high.
Bile The liver secretes bile into the bile duct, which drains into the upper intestine (duodenum). Between meals bile collects in the gallbladder, an expansible storage sac that releases bile when stimulated by the presence of fatty food in the duodenum. Bile contains water, bile salts, and pigments, but no enzymes. Bile salts (mainly sodium taurocholate and sodium glycocholate) are essential for digestion of fats. Fats, because of their tendency to remain in large, waterinsoluble globules, are especially resistant to enzymatic digestion. Bile salts reduce surface tension of fat globules, allowing the churning action of the intestine to break fats into tiny droplets (emulsification). With total surface exposure of fat particles greatly increased, fat-splitting lipases are able to reach and hydrolyze the triglyceride molecules. The yellow-green color of bile is produced by bile pigments, breakdown products of hemoglobin from worn-out red blood cells. Bile pigments also give the feces its characteristic color.
Bile production is only one of the liver’s many functions. This highly versatile organ is a storehouse for glycogen, production center for plasma proteins, site of protein synthesis and detoxification of protein wastes, site for destruction of worn-out red blood cells, and center for metabolism of fat, amino acids, and carbohydrates.
Absorption
Little food is absorbed in the stomach because digestion is still incomplete and because of limited absorptive surface area. However, some materials, such as drugs and alcohol, are absorbed mostly there, which contributes to their rapid action. Most digested food is absorbed from the small intestine where the numerous finger-shaped villi provide an enormous surface area through which materials can pass from the intestinal lumen into the circulation.
Carbohydrates are absorbed almost exclusively as simple sugars (monosaccharides, for example, glucose, fructose, and galactose) because the intestine is virtually impermeable to polysaccharides. Proteins are absorbed principally as their amino acid subunits, although a limited amount of small proteins or peptide fragments sometimes may be absorbed. Both active and passive processes transfer simple sugars and amino acids across the intestinal epithelium.
Immediately after a meal these materials are in such high concentration in the gut that they readily diffuse into the blood, where their concentration is initially lower. However, if absorption were passive only, we would expect transfer to cease as soon as concentrations of a substance became equal on both sides of the intestinal epithelium. Passive transfer alone would permit valuable nutrients to be lost in the feces. In fact, very little is lost because passive transfer is supplemented by an active transport mechanism located in the epithelial cells that transfers food molecules into the blood. Materials thus are moved against their concentration gradient, a process requiring expenditure of energy. Although not all food products are actively transported, those that are, such as glucose, galactose, and most amino acids, are handled by transport mechanisms that are specific for each kind of molecule.
As mentioned previously, fat droplets are emulsified by bile salts and then digested by pancreatic lipase. Triglycerides are broken into fatty acids and monoglycerides, which complex with bile salts to form minute droplets called micelles. When micelles contact the microvilli of the intestinal epithelium, the fatty acids and monoglycerides are absorbed by simple diffusion. They then enter the endoplasmic reticulum of the absorptive cells, where they are resynthesized into triglycerides before passing into lacteals (Figure 34-13B). From the lacteals, fat droplets enter the lymph system (Figure 33-18,) and eventually pass into the blood circulation through the thoracic duct. After a fatty meal, even a peanut butter sandwich, the presence of numerous fat droplets in the blood imparts a milky appearance to the blood plasma.
The importance of the intestine varies widely among animal groups. In invertebrates that have extensive digestive diverticula in which food is broken down and phagocytized, the intestine may serve only as a pathway for conducting wastes out of the body. In other invertebrates with simple stomachs, and in all vertebrates, the intestine is equipped for both digestion and absorption.
Devices for increasing the internal surface area of the intestine are highly developed in vertebrates, but are generally absent among invertebrates. Perhaps the most direct way to increase the absorptive surface of the gut is to increase its length. Coiling of the intestine is common among all vertebrate groups and reaches its highest development in mammals, in which the length of the intestine may exceed eight times the length of the body. Although a coiled intestine is rare among invertebrates, other strategies for increasing surface sometimes occur. For example, the typhlosole of terrestrial oligochaete worms (see Figure 17-12), an inward folding of the dorsal intestinal wall that runs the full length of the intestine, effectively increases internal surface area of the gut in a narrow body lacking space for a coiled intestine.
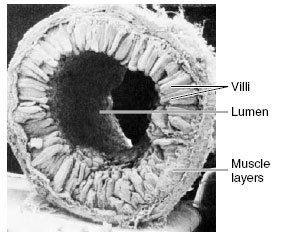 |
| Figure 34-12 Scanning electron micrograph of a rat intestine showing the numerous fingerlike villi that project into the lumen and vastly increase the effective absorptive and secretory surface of the intestine. (×21) |
Lampreys and sharks have longitudinal or spiral folds in their intestine. Other vertebrates have developed elaborate folds (amphibians and reptiles) and minute fingerlike projections called villi (birds and mammals), which give the inner surface of fresh intestinal tissue the appearance of velvet (Figure 34-12). The electron microscope reveals that each cell lining the intestinal cavity additionally is bordered by hundreds of short, delicate processes called microvilli (Figure 34-13C and D). These processes, together with larger villi and intestinal folds, may increase the internal surface area of the intestine more than a million times as compared to a smooth cylinder of the same diameter. This elaborate surface greatly facilitates the absorption of food molecules.
Digestion in the Vertebrate Small Intestine
Food is released into the small intestine through the pyloric sphincter, which relaxes at intervals to allow entry of acidic stomach contents into the initial segment of the small intestine, the duodenum. Two secretions pour into this region: pancreatic juice and bile (Figure 34-14). Both of these secretions have a high bicarbonate content, especially pancreatic juice, which effectively neutralizes gastric acid, raising the pH of the liquefied food mass, now called chyme, from 1.5 to 7 as it enters the duodenum. This change in pH is essential because all intestinal enzymes are effective only in a neutral or slightly alkaline medium.
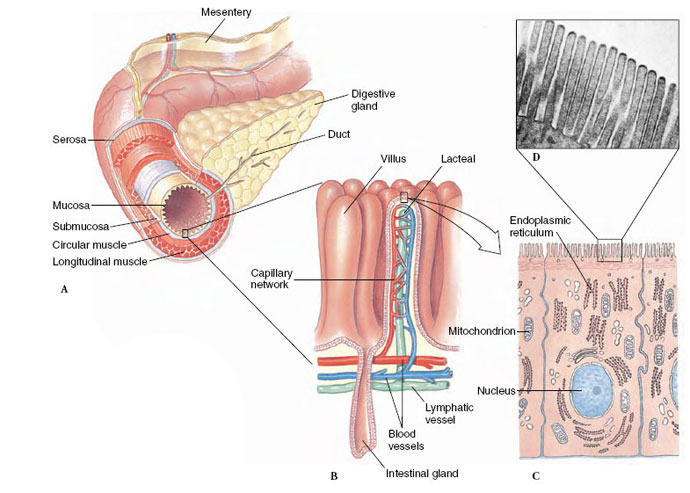 |
| Figure 34-13 Organization of the vertebrate digestive tract, showing A, the successive layers of mucosa, submucosa, muscle, and enveloping serosa; an enzyme-secreting digestive gland (e.g., pancreas); and the thin mesentery that positions the intestine within the body cavity. B, Portion of mucosal lining of intestine, showing fingerlike villi. C, Section of a single cell from the mucosal lining. D, Microvilli on surface of mucosal cell, rat intestine. (×16,400) |
Cells of the intestinal mucosa, like those of the stomach mucosa, are subjected to considerable wear and are constantly undergoing replacement. Cells deep in the crypt between adjacent villi divide rapidly and migrate up the villus. In mammals the cells reach the tip of the villus in about twodays. There they are shed, along with their membrane enzymes, into the lumen at the rate of some 17 billion a day along the length of the human intestine. Before they are shed, however, these cells differentiate into absorptive cells that transport nutrients into the network of blood and lymph vessels, once digestion is complete.
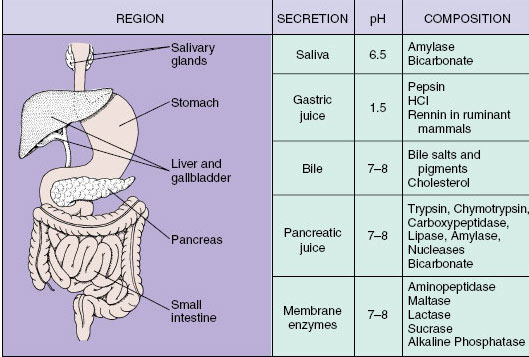 |
| Figure 34-14 Secretions of the mammalian alimentary canal with the principal components and the pH of each secretion. |
Pancreatic Enzymes The pancreatic secretion of vertebrates contains several enzymes of major importance in digestion (Figure 34-14). Two powerful proteases, trypsin and chymotrypsin, continue enzymatic digestion of proteins begun by pepsin, which is now inactivated by the alkalinity of the intestine. Trypsin and chymotrypsin, like pepsin, are highly specific proteases that split apart peptide bonds deep inside the protein molecule. The hydrolysis of a peptide linkage may be shown as: Pancreatic juice also contains carboxypeptidase, which removes amino acids from the carboxyl ends of polypeptides; pancreatic lipase, which hydrolyzes fats into fatty acids and glycerol; pancreatic amylase, a starch-splitting enzyme identical to salivary amylase in its action; and nucleases, which degrade RNA and DNA to nucleotides.
Membrane Enzymes The cells lining the intestine have digestive enzymes embedded in their surface membrane that continue digestion of carbohydrates, proteins, and phosphate compounds (Figure 34-14). These enzymes of the microvillus membrane (Figure 34-13D) include aminopeptidase that splits terminal amino acids from the amino end of short peptides, and several disaccharidases, enzymes that split 12-carbon sugar molecules into 6-carbon units. The disaccharidases include maltase, which splits maltose into two molecules of glucose; sucrase, which splits sucrose to fructose and glucose; and lactase, which breaks lactose (milk sugar) into glucose and galactose. Also present is alkaline phosphatase, an enzyme that attacks a variety of phosphate compounds.
Although milk is the universal food of newborn mammals and one of the most complete human foods,many adult humans cannot digest milk because they are deficient in lactase, the enzyme that hydrolyzes lactose (milk sugar). Lactose intolerance is genetically determined. It is characterized by abdominal bloating, cramps, flatulence, and watery diarrhea, all appearing within 30 to 90 minutes after ingesting milk or its unfermented by-products. (Fermented dairy products, such as yogurt and cheese, create no intolerance problems.)
Northern Europeans and their descendants, which include the majority of North American whites, are most tolerant of milk. Many other ethnic groups are generally intolerant to lactose, including the Japanese, Chinese, Jews in Israel, Eskimos, South American Indians, and most African blacks. Only about 30% of North American blacks are tolerant; those who are tolerant are mostly descendants of slaves brought from east and central Africa where dairying is traditional and tolerance to lactose is high.
Bile The liver secretes bile into the bile duct, which drains into the upper intestine (duodenum). Between meals bile collects in the gallbladder, an expansible storage sac that releases bile when stimulated by the presence of fatty food in the duodenum. Bile contains water, bile salts, and pigments, but no enzymes. Bile salts (mainly sodium taurocholate and sodium glycocholate) are essential for digestion of fats. Fats, because of their tendency to remain in large, waterinsoluble globules, are especially resistant to enzymatic digestion. Bile salts reduce surface tension of fat globules, allowing the churning action of the intestine to break fats into tiny droplets (emulsification). With total surface exposure of fat particles greatly increased, fat-splitting lipases are able to reach and hydrolyze the triglyceride molecules. The yellow-green color of bile is produced by bile pigments, breakdown products of hemoglobin from worn-out red blood cells. Bile pigments also give the feces its characteristic color.
Bile production is only one of the liver’s many functions. This highly versatile organ is a storehouse for glycogen, production center for plasma proteins, site of protein synthesis and detoxification of protein wastes, site for destruction of worn-out red blood cells, and center for metabolism of fat, amino acids, and carbohydrates.
Absorption
Little food is absorbed in the stomach because digestion is still incomplete and because of limited absorptive surface area. However, some materials, such as drugs and alcohol, are absorbed mostly there, which contributes to their rapid action. Most digested food is absorbed from the small intestine where the numerous finger-shaped villi provide an enormous surface area through which materials can pass from the intestinal lumen into the circulation.
Carbohydrates are absorbed almost exclusively as simple sugars (monosaccharides, for example, glucose, fructose, and galactose) because the intestine is virtually impermeable to polysaccharides. Proteins are absorbed principally as their amino acid subunits, although a limited amount of small proteins or peptide fragments sometimes may be absorbed. Both active and passive processes transfer simple sugars and amino acids across the intestinal epithelium.
Immediately after a meal these materials are in such high concentration in the gut that they readily diffuse into the blood, where their concentration is initially lower. However, if absorption were passive only, we would expect transfer to cease as soon as concentrations of a substance became equal on both sides of the intestinal epithelium. Passive transfer alone would permit valuable nutrients to be lost in the feces. In fact, very little is lost because passive transfer is supplemented by an active transport mechanism located in the epithelial cells that transfers food molecules into the blood. Materials thus are moved against their concentration gradient, a process requiring expenditure of energy. Although not all food products are actively transported, those that are, such as glucose, galactose, and most amino acids, are handled by transport mechanisms that are specific for each kind of molecule.
As mentioned previously, fat droplets are emulsified by bile salts and then digested by pancreatic lipase. Triglycerides are broken into fatty acids and monoglycerides, which complex with bile salts to form minute droplets called micelles. When micelles contact the microvilli of the intestinal epithelium, the fatty acids and monoglycerides are absorbed by simple diffusion. They then enter the endoplasmic reticulum of the absorptive cells, where they are resynthesized into triglycerides before passing into lacteals (Figure 34-13B). From the lacteals, fat droplets enter the lymph system (Figure 33-18,) and eventually pass into the blood circulation through the thoracic duct. After a fatty meal, even a peanut butter sandwich, the presence of numerous fat droplets in the blood imparts a milky appearance to the blood plasma.
Support our developers




