Characteristics and Natural History of Reptilian Orders
Characteristics and
Natural History of
Reptilian Orders
Anapsid Reptiles: Subclass Anapsida
Order Testudines (Chelonia): Turtles
Turtles descended from one of the earliest anapsid lineages, probably a group known as the procolophonids of the late Permian, but turtles themselves do not appear in the fossil record until the Upper Triassic, some 200 million years ago. From the Triassic, turtles plodded on to the present with very little change to their early morphology. They are enclosed in shells consisting of a dorsal carapace (Fr., from Sp., carapacho, covering) and a ventral plastron (Fr., breastplate). The shell is composed of two layers: an outer horny layer of keratin and an inner layer of bone. New layers of keratin are laid down beneath the old as the turtle grows and ages. The bony layer is a fusion of ribs, vertebrae, and many dermally-ossifying elements (Figure 28-6). Unique among vertebrates, turtle limbs and limb girdles are located inside the ribs! Lacking teeth, the turtle jaw is provided with tough, horny plates for gripping food (Figure 28-7).
Clumsy and unlikely as they appear to be within their protective shells, turtles are nonetheless a varied and ecologically diverse group that seems able to adjust to human presence.
One consequence of living in a rigid shell with fused ribs is that a turtle cannot expand its chest to breathe. Turtles solved this problem by employing certain abdominal and pectoral muscles as a “diaphragm.” Air is drawn in by increasing abdominal cavity volume by contracting limb flank muscles. Exhalation is also active and is accomplished by drawing the shoulder girdle back into the shell, thus compressing the viscera and forcing air out of the lungs. Breathing is visible as the bellowslike movements of the turtle’s “limb pockets”: folds of skin between the limbs and the rigid shell. The movements of the limbs during walking also help to ventilate the lungs. Many aquatic turtles gain enough oxygen by just pumping water in and out of a vascularized mouth cavity; this activity enables them to remain submerged for long periods when inactive. When active they must lungbreathe more frequently.
A turtle’s brain, like that of other reptiles, is small, never exceeding 1% of body weight. The cerebrum, however, is larger than that of an amphibian, and turtles are able to learn a maze about as quickly as a rat. Turtles have both a middle and an inner ear, but perception of sound is poor. Not unexpectedly, therefore, turtles are virtually mute (the biblical “voice of the turtle” refers to the turtledove, a bird), although many tortoises utter grunting or roaring sounds during mating (Figure 28-8). Compensating for poor hearing are a good sense of smell, acute vision, and color perception evidently as good as that of humans.
Turtles are oviparous. Fertilization is internal and all turtles, even marine forms, bury their shelled, amniotic eggs in the ground. Usually considerable care is exercised in constructing the nest, but once eggs are deposited and covered, the female deserts them. An odd feature of turtle reproduction is that in some turtle families, as in all crocodilians and some lizards, nest temperature determines the sex of the hatchlings. In turtles, low temperatures during incubation produce males and high temperatures produce females. All temperature-dependent reptiles lack sex chromosomes.

The great marine turtles, buoyed
by their aquatic environment, may reach
2 m in length and 725 kg in weight. One such heavyweight is the leatherback.
The green turtle (Figure 28-9), so
named because of its greenish body fat,
may exceed 360 kg, although most individuals
of this economically valuable
and heavily exploited species seldom
live long enough to reach anything
approaching this size. Some land tortoises
may weigh several hundred kilograms,
such as the giant tortoises of the
Galápagos Islands that so intrigued
Darwin during his visit there in 1835.
Most tortoises are rather slow moving;
an hour of determined trudging carries
a large Galápagos tortoise approximately
300 m (although they may move
much more rapidly for short distances).
Their low metabolism probably
explains their longevity, for some are
believed to live more than 150 years.
The shell, like a medieval coat of armor, offers obvious advantages. The head and appendages can be drawn in for protection. The familiar box tortoise (Terrapene carolina) has a plastron that is hinged, forming two movable parts that can be pulled up against the carapace so tightly that one can hardly force a knife blade between the shells. Some turtles, such as the large eastern snapping turtle (Chelydra serpentina), have reduced shells, making complete withdrawal for protection quite impossible. Snappers, however, have another formidable defense, as their name implies (Figure 28-7). They are entirely carnivorous, living on fish, frogs, waterfowl, or almost anything that comes within reach of their powerful jaws. The alligator snapper lures unwary fish into its mouth with a “bait” (Figure 28-10). Snappers are wholly aquatic and come ashore only to lay their eggs.
Diapsid Reptiles: Subclass Diapsida
Diapsid reptiles, those reptiles having a skull with two pairs of temporal openings (Figure 28-2), are classified into three lineages. The two with living representatives are the superorder Lepidosauria, containing lizards, snakes, worm lizards, and Sphenodon; and the superorder Archosauria, containing the crocodilians.
Order Squamata: Lizards, Snakes, and Worm Lizards
The squamates are the most recent and diverse products of diapsid evolution, comprising approximately 95% of all known living reptiles. Lizards appeared in the fossil record as early as the Permian, but they did not begin their radiation until the Cretaceous period of the Mesozoic era when the dinosaurs were at the climax of their radiation. Snakes appeared during the late Cretaceous period, probably from a group of lizards whose descendants include the Gila monster and monitor lizards. Two specializations in particular characterize snakes: extreme elongation of the body and accompanying displacement and rearrangement of internal organs; and specializations for eating large prey. The amphisbaenians (worm lizards), which first appear in the fossil record of the early Cenozoic era, have structural specializations associated with a burrowing habit.
The diapsid skulls of squamates
are modified from the ancestral diapsid
condition by loss of dermal bone ventral
and posterior to the lower temporal
opening. This modification has
allowed evolution in most lizards and
snakes of a mobile skull having movable
joints. Such a skull is called a kinetic skull. The quadrate, which in other reptiles is fused to the skull,
has a joint at its dorsal end, as well as
its usual articulation with the lower
jaw. In addition, there are joints in
the palate and across the roof of the
skull that allow the snout to be tilted
upward (Figure 28-11). The specialized
mobility of the skull enables squamates
to seize and manipulate their
prey, it also increases the effective
closing force of the jaw musculature.
The skull of snakes is even more kinetic
than that of lizards. Such exceptional
skull mobility is considered a
major factor in the diversification of
lizards and snakes.
Suborder Sauria: Lizards - Lizards are an extremely diverse group, including terrestrial, burrowing, aquatic, arboreal and aerial members. Among the more familiar groups in this varied suborder are geckos (Figure 28-12), small, agile, mostly nocturnal forms with adhesive toe pads that enable them to walk upside down and on vertical surfaces; iguanids, including most of the familiar New World lizards, often brightly colored with ornamental crests, frills, and throat fans, and a group that includes the remarkable marine iguana of the Galápagos Islands (Figure 28-13); skinks, with elongate bodies and reduced limbs in many species; and chameleons, a group of arboreal lizards, mostly of Africa and Madagascar. Chameleons are entertaining creatures that catch insects with the stickytipped tongue that can be flicked accurately and rapidly to a distance greater than the length of their body (Figure 28-
14). The great majority of
lizards have four limbs and relatively
short bodies, but in many the limbs are
degenerate, and a few such as the glass
lizards (Figure 28-15) are completely
limbless.
Most lizards have movable eyelids, whereas a snake’s eyes are permanently covered with a transparent cap. Lizards have keen vision for daylight (retinas rich in both cones and rods), although one group, the nocturnal geckos, has retinas composed entirely of rods. Most lizards have an external ear that snakes lack. The inner ear of lizards is variable in structure, but as with other reptiles, hearing does not play an important role in the lives of most lizards. Geckos are exceptions because males are strongly vocal (to announce territory and discourage the approach of other males, and they must, of course, hear their own vocalizations. Other species of lizards vocalize in defensive behavior.
Many lizards live in the world’s hot and arid regions. Since their skin lacks glands, water loss by this avenue is much reduced. They produce a semisolid urine with a high content of crystalline uric acid. This is an excellent mechanism for conserving water and is found in other groups living successfully in arid habitats (birds, insects, and pulmonate snails). Some, such as the Gila monster of southwestern United States deserts, store fat in their tails, which they use during drought to provide energy and metabolic water (Figure 28-16). The Gila monster, and its close relative, the bearded lizard, are the only lizards capable of delivering a venomous bite. The way many lizards keep their body temperature relatively constant by behavioral thermoregulation is described in Homeostasis.
Suborder Amphisbaenia: Worm
Lizards - The somewhat inappropriate
common name “worm lizards”
describes a group of highly specialized,
burrowing forms that are neither
worms nor true lizards but certainly
are related to the latter. The name of
the suborder literally means “double
walk,” in reference to their peculiar
ability to move backward nearly
as effectively as forward. They have
elongate, cylindrical bodies of nearly uniform diameter, and most lack any
trace of external limbs (Figure 28-17).
The soft skin is divided into numerous
rings, which combined with absence
of visible eyes and ears (both are hidden
under skin) make amphisbaenians
look like earthworms. The resemblance,
although superficial, is the
kind of structural convergence that
often occurs when two unrelated
groups come to occupy similar habitats.
Amphisbaenians have an extensive
distribution in South America and
tropical Africa. In the United States,
one species, Rhineura florida, is
found in Florida where it is known as
the “graveyard snake.”
Suborder Serpentes: Snakes - Snakes are entirely limbless and lack both pectoral and pelvic girdles (the latter persists as a vestige in pythons, boas, and some other snakes). The numerous vertebrae of snakes, shorter and wider than those of tetrapods, permit quick lateral undulations through grass and over rough terrain. The ribs increase rigidity of the vertebral column, providing more resistance to lateral stresses. The elevation of the neural spine gives the numerous muscles more leverage.
The highly kinetic skull and feeding apparatus of snakes, which enable them to eat prey several times their own diameter, are perhaps their most remarkable specialization. The two halves of the lower jaw (mandibles) are joined only by muscles and skin, allowing them to spread widely apart. Many of the skull bones are so loosely articulated that the entire skull can flex asymmetrically to accommodate oversized prey (Figures 28-11 and 28-18). Since the snake must keep breathing during the slow process of swallowing, the tracheal opening (glottis) is thrust forward between the two mandibles.
The cornea of the snake’s eye is permanently protected with a transparent membrane called a spectacle, which, together with reduced eyeball mobility, gives snakes the cold, unblinking stare that many people find unnerving. Most snakes have relatively poor vision, arboreal snakes of the tropical forest being a conspicuous exception (Figure 28-19). Some arboreal snakes possess excellent binocular vision that helps them track prey through branches where scent trails would be impossible to follow.
Snakes have no external ears or tympanic membrane. This condition, together with the absence of any obvious response to aerial sounds, led to the widespread opinion that snakes are totally deaf. But snakes do have internal ears, and recent work has shown quite clearly that within a limited range of low frequencies (100 to 700 Hz), hearing in snakes compares favorably with that of most lizards. Snakes are also quite sensitive to vibrations carried in the ground.
Nevertheless, for most snakes it is the chemical senses and not vision and hearing that are employed to hunt their prey. In addition to the usual olfactory areas in the nose, which are not well developed, there are Jacobson’s organs, a pair of pitlike organs in the roof of the mouth. These are lined with an olfactory epithelium and are richly innervated. The forked tongue, flicking through the air, picks up scent particles and conveys them to the mouth; the tongue is then drawn past Jacobson’s organs and information is then transmitted to the brain where scents are identified (Figure 28-20).
As with lizards, the snake’s body is entirely covered with a tough, impervious skin. The hard scales are set together, sometimes overlapping like shingles on a roof, with the skin folded inward between the scales. The skin is not elastic, and when required to stretch, as it must after the snake has enjoyed a hefty meal, it does so in a novel manner. The skin folds are pulled out straight, leaving the scales separated like islands on the skin.
Snakes have evolved several solutions to the obvious problem of movement without legs. The most typical pattern of movement is lateral undulation (Figure 28-21A). Movement follows an S-shaped path, with the snake propelling itself by exerting lateral force against surface irregularities. The snake seems to “flow,” since the moving loops appear stationary with respect to the ground. Lateral undulatory movement is fast and efficient under most but not all circumstances. Concertina movement (Figure 28-21B) enables a snake to move in a narrow passage, as when climbing a tree by using irregular channels in the bark. The snake extends forward while bracing S-shaped loops against the sides of the channel. To advance in a straight line as when stalking prey, many heavy-bodied snakes employ rectilinear movement. Two or three sections of the body rest on the ground to support the snake’s weight. Intervening sections are lifted free of the ground and pulled forward by muscles (shown in red in Figure 28-21C) that originate on ribs and insert on the ventral skin. Rectilinear movement is a slow but effective way of moving inconspicuously toward prey, even when there are no surface irregularities. Side-winding is a fourth form of movement that enables desert vipers to move with surprising speed across loose, sandy surfaces with minimum surface contact. The sidewinder rattlesnake moves by throwing its body forward in loops with its body lying at an angle of about 60 degrees to its direction of travel.
Snakes of the subfamily Crotalinae
within the family Viperidae are
called pit vipers because they possess
special heat-sensitive pit organs
on their heads, located between the
nostrils and the eyes (Figures 28-20
and 28-22). All of the best-known
North American venomous snakes are
pit vipers, such as the several species
of rattlesnakes, the cottonmouth, and
the copperhead. The pits are supplied with a dense packing of free nerve
endings from the fifth cranial nerve.
These respond to radiant energy in
the long-wave infrared (5000 to
15,000 nm) and are especially sensitive
to heat emitted by warm-bodied
birds and mammals that are their
food (infrared wavelengths of about
10,000 nm). Some measurements suggest
that pit organs can distinguish
temperature differences of only
0.003° C from a radiating surface. Pit
vipers use pit organs to track warmblooded
prey and to aim strikes with
great accuracy, as effectively in total
darkness as in daylight. Boa constrictors
and pythons also have heat
receptors (in their lips), but the
anatomy is quite different from that of
pit vipers, suggesting that they probably
evolved independently.
All vipers have a pair of teeth on
the maxillary bones modified as fangs.
These lie in a membrane sheath when
the mouth is closed. When the viper
strikes, a special muscle and bone
lever system erects the fangs when the
mouth opens (Figure 28-23). The fangs
are driven into the prey by the thrust
of the strike, and venom is injected
into the wound through a channel in
the fangs. A viper immediately releases
its prey after the bite and waits until it
is paralyzed or dead. Then the snake
swallows the prey whole. The bite of a
pit viper can be dangerous to humans,
although in many instances the snake
injects very little venom when it bites.
Approximately 8000 bites but only 12
deaths from pit vipers are reported
each year in the United States.
The tropical and subtropical countries are the homes of most species of snakes, both of the venomous and nonvenomous varieties. Even in these countries, less than one-third of snakes are venomous. Nonvenomous snakes kill their prey by constriction (Figure 28-24) or by biting and swallowing. Their diet tends to be restricted, many feeding principally on rodents, whereas others feed on fishes, frogs, and insects. Some African, Indian, and neotropical snakes are egg eaters.
Venomous snakes are usually
divided into four groups based on the
type of fangs. Vipers (family Viperidae)
have highly developed tubular fangs at
the front of the mouth; the group
includes the American pit vipers previously
mentioned and Old World true
vipers, which lack facial heat-sensing
pits. Among the latter are the common
European adder and the African puff
adder. A second family of venomous
snakes (family Elapidae) has short, permanently
erect fangs. In this group are
the cobras (Figure 28-25), mambas,
coral snakes, and kraits. The highly
venomous sea snakes are usually
placed in a third family (Hydrophiidae). The very large family Colubridae, which
contains most of the familiar (and nonvenomous)
snakes, does include at least
several snakes that have been responsible
for human fatalities. Two examples
are the African boomslang and the
African twig snake, both rear-fanged
snakes that normally use their venom to
quiet struggling prey.
Even the saliva of harmless snakes possesses limited toxic qualities, and it is logical that there was a natural selection for this toxic tendency as snakes evolved. Snake venoms have traditionally been divided into two types. The neurotoxic type acts mainly on the nervous system, affecting the optic nerves (causing blindness) or the phrenic nerve of the diaphragm (causing paralysis of respiration). The hemorrhagin type breaks down red blood corpuscles and blood vessels and produces extensive hemorrhaging of blood into tissue spaces. In fact, most snake venoms are complex mixtures of various fractions that attack different organs in specific ways; they seldom can be assigned categorically to one or the other of the traditional types.
The toxicity of a venom is measured by the median lethal dose on laboratory animals (LD50). By this standard the venoms of the Australian tiger snake and some of the sea snakes appear to be the most deadly of venomous drop for drop. However, several larger snakes are more dangerous. The aggressive king cobra, which may exceed 5.5 m in length, is the largest and probably the most dangerous of all poisonous snakes. In India and Burma, where snakes come in frequent contact with poorly shod people, some 200,000 snake bites cause more than 10,000 deaths each year. Russell’s viper is responsible for some 1000 deaths each year in Burma alone. The world total for deaths from snakebite is estimated at 25,000 each year.
Most snakes are oviparous (L. ovum, egg, + parere, to bring forth) species that lay their shelled, elliptical eggs beneath rotten logs, under rocks, or in holes in the ground. Most of the remainder, including all the American pit vipers, except the tropical bushmaster, are ovoviviparous (L. ovum, egg, + vivus, living, + parere, to bring forth), giving birth to well-formed young. Very few snakes are viviparous (L. vivus, living, + parere, to bring forth); in these snakes a primitive placenta forms, permitting exchange of materials between the embryonic and maternal bloodstreams. Snakes are able to store sperm and can lay several clutches of fertile eggs at long intervals after a single mating.
Order Sphenodonta: The Tuatara
The order Sphenodonta is represented by two living species of the genus Sphenodon (Gr. sphenos, wedge, + odontos, tooth) of New Zealand (Figure 28-26). The tuatara is the sole survivor of the sphenodontid lineage that radiated modestly during the early Mesozoic era but declined toward the end of the Mesozoic. Several species of the tuatara were once widespread throughout the two main islands of New Zealand but the two living species are now restricted to small islets of Cook Strait and off the northeast coast of North Island. On some of these islands, under protection by the New Zealand government, they are prospering.
The tuatara is a lizardlike form 66 cm long or less that lives in burrows often shared with petrels. They are slow-growing animals with a long life; one is recorded to have lived 77 years.
The tuatara has captured the interest of zoologists because of numerous features that are almost identical to those of Mesozoic fossils 200 million years ago. These features include a diapsid skull with two temporal openings bounded by complete arches. Tuataras also bear a welldeveloped median parietal eye complete with elements of cornea, lens, and retina (although since it is buried beneath opaque skin this “third eye” can register only changes in light intensity, and its function, if any, remains unknown). In many other respects Sphenodon resembles lizards of the early Mesozoic. Sphenodon represents one of the slowest rates of evolution known among vertebrates.
Order Crocodilia: Crocodiles and Alligators
The modern crocodilians are the only surviving reptiles of the archosaurian lineage that gave rise to the great Mesozoic radiation of dinosaurs and their kin and to the birds. Although modern crocodiles belong to a lineage that began its radiation in the late Cretaceous period, they differ little in structural details from primitive crocodilians of the early Mesozoic. Having remained mostly unchanged for nearly 200 million years, crocodilians face an uncertain future in a world dominated by humans. Modern crocodilians are divided into three families: alligators and caimans, mostly a New World group; crocodiles, which are widely distributed and include the saltwater crocodile, one of the largest living reptiles; and gavials, represented by a single species in India and Burma.
All crocodilians have an elongate, robust, well-reinforced skull and massive jaw musculature arranged to provide a wide gape and rapid, powerful closure. Teeth are set in sockets, a type of dentition called thecodont that was typical of all archosaurs as well as the earliest birds. Another adaptation, found in no other vertebrate except mammals, is a complete secondary palate. This innovation allows crocodilians to breathe when the mouth is filled with water or food (or both). Crocodilians also share a four-chambered heart with birds and mammals.
The estuarine crocodile (Crocodylus porosus), found in southern Asia, and the Nile crocodile (C. niloticus; Figure 28-27A) grow to great size (adults weighing 1000 kg have been reported) and are swift and aggressive. Crocodiles are known to attack animals as large as cattle, deer, and people. Alligators (Figure 28-27B) are usually less aggressive than crocodiles and far less dangerous to humans. Large alligators are powerful animals nevertheless, and adults have almost no enemies but humans. The chink in their formidable armor is the developmental stages. Nests left unguarded by the mother are almost certain to be discovered and raided by any of several mammals that relish eggs, and the young hatchlings may be devoured by large fish.
Alligators are able to make definite vocalizations. The male alligator can give loud bellows in the mating season. In the United States, Alligator mississipiensis (Figure 28-27B) is the only species of alligator; Crocodylus acutus, restricted to extreme southern Florida, is the only species of crocodile.
Alligators and crocodiles are oviparous. Usually 20 to 50 eggs are laid in a mass of dead vegetation and guarded by the mother. The mother hears vocalizations from the hatching young and responds by opening the nest to allow the hatchlings to escape. As with many turtles and some lizards, incubation temperature of the eggs determines sex ratio of the offspring. However, unlike turtles (low nest temperatures produce only females, whereas high nest temperatures produce only males. This results in highly unbalanced sex ratios in some areas. For example, in one study area in Louisiana, female hatchlings outnumbered males five to one.
Crocodiles and alligators can be distinguished on the basis of head morphology. Crocodiles have a relatively narrow snout, and when their mouths are closed, the fourth lower jaw tooth is visible. Alligators generally have a broader snout, and their fourth lower jaw tooth is hidden by fitting into a notch in the upper jaw (Figure 28-27). Gavials have very narrow snouts, and are largely fish eaters.
Anapsid Reptiles: Subclass Anapsida
Order Testudines (Chelonia): Turtles
Turtles descended from one of the earliest anapsid lineages, probably a group known as the procolophonids of the late Permian, but turtles themselves do not appear in the fossil record until the Upper Triassic, some 200 million years ago. From the Triassic, turtles plodded on to the present with very little change to their early morphology. They are enclosed in shells consisting of a dorsal carapace (Fr., from Sp., carapacho, covering) and a ventral plastron (Fr., breastplate). The shell is composed of two layers: an outer horny layer of keratin and an inner layer of bone. New layers of keratin are laid down beneath the old as the turtle grows and ages. The bony layer is a fusion of ribs, vertebrae, and many dermally-ossifying elements (Figure 28-6). Unique among vertebrates, turtle limbs and limb girdles are located inside the ribs! Lacking teeth, the turtle jaw is provided with tough, horny plates for gripping food (Figure 28-7).
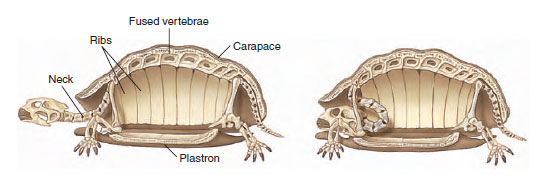 |
| Figure 28-6 Skeleton and shell of a turtle, showing fusion of vertebrae and ribs with the carapace. The long and flexible neck allows the turtle to withdraw its head into its shell for protection. |
 |
| Figure 28-7 Snapping turtle, Chelydra serpentina, showing the absence of teeth. Instead, the jaw edges are covered with a horny plate. |
Clumsy and unlikely as they appear to be within their protective shells, turtles are nonetheless a varied and ecologically diverse group that seems able to adjust to human presence.
One consequence of living in a rigid shell with fused ribs is that a turtle cannot expand its chest to breathe. Turtles solved this problem by employing certain abdominal and pectoral muscles as a “diaphragm.” Air is drawn in by increasing abdominal cavity volume by contracting limb flank muscles. Exhalation is also active and is accomplished by drawing the shoulder girdle back into the shell, thus compressing the viscera and forcing air out of the lungs. Breathing is visible as the bellowslike movements of the turtle’s “limb pockets”: folds of skin between the limbs and the rigid shell. The movements of the limbs during walking also help to ventilate the lungs. Many aquatic turtles gain enough oxygen by just pumping water in and out of a vascularized mouth cavity; this activity enables them to remain submerged for long periods when inactive. When active they must lungbreathe more frequently.
 |
| Figure 28-8 Mating Galápagos tortoises. The male has a concave plastron that fits over the highly convex carapace of the female, helping to provide stability during mating. Males utter a roaring sound during mating, the only time they are known to emit vocalizations |
A turtle’s brain, like that of other reptiles, is small, never exceeding 1% of body weight. The cerebrum, however, is larger than that of an amphibian, and turtles are able to learn a maze about as quickly as a rat. Turtles have both a middle and an inner ear, but perception of sound is poor. Not unexpectedly, therefore, turtles are virtually mute (the biblical “voice of the turtle” refers to the turtledove, a bird), although many tortoises utter grunting or roaring sounds during mating (Figure 28-8). Compensating for poor hearing are a good sense of smell, acute vision, and color perception evidently as good as that of humans.
Turtles are oviparous. Fertilization is internal and all turtles, even marine forms, bury their shelled, amniotic eggs in the ground. Usually considerable care is exercised in constructing the nest, but once eggs are deposited and covered, the female deserts them. An odd feature of turtle reproduction is that in some turtle families, as in all crocodilians and some lizards, nest temperature determines the sex of the hatchlings. In turtles, low temperatures during incubation produce males and high temperatures produce females. All temperature-dependent reptiles lack sex chromosomes.
| Figure 28-9 Green sea turtle, Chelonia mydas. Green turtles are herbivores that subsist on marine grasses and algae. Sea turtles range widely in the oceans, returning to land only to deposit their eggs. Sea turtles are found in all tropical oceans. |
The shell, like a medieval coat of armor, offers obvious advantages. The head and appendages can be drawn in for protection. The familiar box tortoise (Terrapene carolina) has a plastron that is hinged, forming two movable parts that can be pulled up against the carapace so tightly that one can hardly force a knife blade between the shells. Some turtles, such as the large eastern snapping turtle (Chelydra serpentina), have reduced shells, making complete withdrawal for protection quite impossible. Snappers, however, have another formidable defense, as their name implies (Figure 28-7). They are entirely carnivorous, living on fish, frogs, waterfowl, or almost anything that comes within reach of their powerful jaws. The alligator snapper lures unwary fish into its mouth with a “bait” (Figure 28-10). Snappers are wholly aquatic and come ashore only to lay their eggs.
 |
| Figure 28-10 Alligator snapping turtle Macroclemys temmincki of the southeastern United States lies on the bottom, mouth agape, luring fish and other unwary prey by undulating a pink, wormlike protrusion from its tongue. Any prey attempting to eat the bait is instantly captured in powerful jaws. |
Diapsid Reptiles: Subclass Diapsida
Diapsid reptiles, those reptiles having a skull with two pairs of temporal openings (Figure 28-2), are classified into three lineages. The two with living representatives are the superorder Lepidosauria, containing lizards, snakes, worm lizards, and Sphenodon; and the superorder Archosauria, containing the crocodilians.
Order Squamata: Lizards, Snakes, and Worm Lizards
The squamates are the most recent and diverse products of diapsid evolution, comprising approximately 95% of all known living reptiles. Lizards appeared in the fossil record as early as the Permian, but they did not begin their radiation until the Cretaceous period of the Mesozoic era when the dinosaurs were at the climax of their radiation. Snakes appeared during the late Cretaceous period, probably from a group of lizards whose descendants include the Gila monster and monitor lizards. Two specializations in particular characterize snakes: extreme elongation of the body and accompanying displacement and rearrangement of internal organs; and specializations for eating large prey. The amphisbaenians (worm lizards), which first appear in the fossil record of the early Cenozoic era, have structural specializations associated with a burrowing habit.
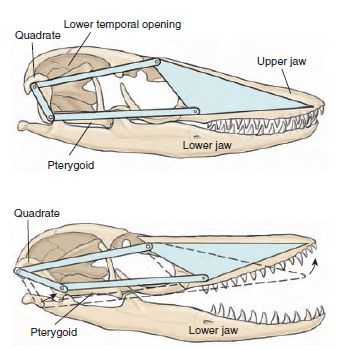 |
| Figure 28-11 Kinetic diapsid skull of a modern lizard (monitor lizard, Varanus sp.) showing the joints that allow the snout and upper jaw to move on the rest of the skull. The quadrate can move at its dorsal end and ventrally at both the lower jaw and the pterygoid. The front part of the braincase is also flexible, allowing the snout to be raised. Note that the lower temporal opening is very large with no lower border; this modification of the diapsid condition, common in modern lizards, provides space for expansion of large jaw muscles. The upper temporal opening lies dorsal and medial to the postorbitalsquamosal arch and is not visible in this drawing. |
Suborder Sauria: Lizards - Lizards are an extremely diverse group, including terrestrial, burrowing, aquatic, arboreal and aerial members. Among the more familiar groups in this varied suborder are geckos (Figure 28-12), small, agile, mostly nocturnal forms with adhesive toe pads that enable them to walk upside down and on vertical surfaces; iguanids, including most of the familiar New World lizards, often brightly colored with ornamental crests, frills, and throat fans, and a group that includes the remarkable marine iguana of the Galápagos Islands (Figure 28-13); skinks, with elongate bodies and reduced limbs in many species; and chameleons, a group of arboreal lizards, mostly of Africa and Madagascar. Chameleons are entertaining creatures that catch insects with the stickytipped tongue that can be flicked accurately and rapidly to a distance greater than the length of their body (Figure 28-
 |
| Figure 28-12 Tokay, Gekko gecko, of Southeast Asia has a true voice and is named after the strident repeated to-kay, to- kay call. |
 |
| Figure 28-13 A large male marine iguana, Amblyrhynchus cristatus, of the Galápagos Islands, feeding underwater on algae. This is the only marine lizard in the world. It has special salt-removing glands in the eye orbits and long claws that enable it to cling to the bottom while feeding on small red and green algae, its principal diet. It may dive to depths exceeding 10 m (33 feet) and remain submerged more than 30 minutes. |
Most lizards have movable eyelids, whereas a snake’s eyes are permanently covered with a transparent cap. Lizards have keen vision for daylight (retinas rich in both cones and rods), although one group, the nocturnal geckos, has retinas composed entirely of rods. Most lizards have an external ear that snakes lack. The inner ear of lizards is variable in structure, but as with other reptiles, hearing does not play an important role in the lives of most lizards. Geckos are exceptions because males are strongly vocal (to announce territory and discourage the approach of other males, and they must, of course, hear their own vocalizations. Other species of lizards vocalize in defensive behavior.
Many lizards live in the world’s hot and arid regions. Since their skin lacks glands, water loss by this avenue is much reduced. They produce a semisolid urine with a high content of crystalline uric acid. This is an excellent mechanism for conserving water and is found in other groups living successfully in arid habitats (birds, insects, and pulmonate snails). Some, such as the Gila monster of southwestern United States deserts, store fat in their tails, which they use during drought to provide energy and metabolic water (Figure 28-16). The Gila monster, and its close relative, the bearded lizard, are the only lizards capable of delivering a venomous bite. The way many lizards keep their body temperature relatively constant by behavioral thermoregulation is described in Homeostasis.
 |
 |
|
| Figure 28-14 A chameleon snares a dragonfly. After cautiously edging close to its target, the chameleon suddenly lunges forward, anchoring its tail and feet to the branch. A split second later, it launches its stickytipped, foot-long tongue to trap the prey. The eyes of this common European chameleon (Chamaeleo chamaeleon) are swiveled forward to provide binocular vision and excellent depth perception. |
Figure 28-15 A glass lizard, Ophisaurus sp., of the southeastern United States. This legless lizard feels stiff and brittle to the touch and has an extremely long, fragile tail that readily fractures when the animal is struck or seized. Most specimens, such as this one, have only a partly regenerated tip to replace a much longer tail previously lost. Glass lizards can be readily distinguished from snakes by the deep, flexible groove running along each side of the body. They feed on worms, insects, spiders, birds’ eggs, and small reptiles. |
 |
 |
| Figure 28-16 Gila monster, Heloderma suspectum, of southwestern United States desert regions and the related Mexican bearded lizard are the only venomous lizards known. These brightly colored, clumsy-looking lizards feed principally on birds’ eggs, nesting birds, mammals, and insects. Unlike poisonous snakes, the Gila monster secretes venom from glands in its lower jaw. The chewing bite is painful to humans but seldom fatal. |
 |
| Figure 28-17 A worm lizard of the suborder Amphisbaenia. Worm lizards are burrowing forms with a solidly constructed skull used as a digging tool. The species pictured, Amphisbaena alba, is widely distributed in South America. |
Suborder Serpentes: Snakes - Snakes are entirely limbless and lack both pectoral and pelvic girdles (the latter persists as a vestige in pythons, boas, and some other snakes). The numerous vertebrae of snakes, shorter and wider than those of tetrapods, permit quick lateral undulations through grass and over rough terrain. The ribs increase rigidity of the vertebral column, providing more resistance to lateral stresses. The elevation of the neural spine gives the numerous muscles more leverage.
 |
| Figure 28-18 Black rat snake, Elaphe obsoleta obsoleta, swallowing a chipmunk. |
The highly kinetic skull and feeding apparatus of snakes, which enable them to eat prey several times their own diameter, are perhaps their most remarkable specialization. The two halves of the lower jaw (mandibles) are joined only by muscles and skin, allowing them to spread widely apart. Many of the skull bones are so loosely articulated that the entire skull can flex asymmetrically to accommodate oversized prey (Figures 28-11 and 28-18). Since the snake must keep breathing during the slow process of swallowing, the tracheal opening (glottis) is thrust forward between the two mandibles.
The cornea of the snake’s eye is permanently protected with a transparent membrane called a spectacle, which, together with reduced eyeball mobility, gives snakes the cold, unblinking stare that many people find unnerving. Most snakes have relatively poor vision, arboreal snakes of the tropical forest being a conspicuous exception (Figure 28-19). Some arboreal snakes possess excellent binocular vision that helps them track prey through branches where scent trails would be impossible to follow.
 |
| Figure 28-19 Parrot snake, Leptophis ahaetulla. The slender body of this Central American tree snake is an adaptation for sliding along branches without weighing them down. |
Snakes have no external ears or tympanic membrane. This condition, together with the absence of any obvious response to aerial sounds, led to the widespread opinion that snakes are totally deaf. But snakes do have internal ears, and recent work has shown quite clearly that within a limited range of low frequencies (100 to 700 Hz), hearing in snakes compares favorably with that of most lizards. Snakes are also quite sensitive to vibrations carried in the ground.
 |
| Figure 28-20 A blacktail rattlesnake, Crotalus molossus, flicks its tongue to smell its surroundings. Scent particles trapped on the tongue’s surface are transferred to Jacobson’s organs, olfactory organs in the roof of the mouth. Note the heat-sensitive pit organ between the nostril and eye. |
Nevertheless, for most snakes it is the chemical senses and not vision and hearing that are employed to hunt their prey. In addition to the usual olfactory areas in the nose, which are not well developed, there are Jacobson’s organs, a pair of pitlike organs in the roof of the mouth. These are lined with an olfactory epithelium and are richly innervated. The forked tongue, flicking through the air, picks up scent particles and conveys them to the mouth; the tongue is then drawn past Jacobson’s organs and information is then transmitted to the brain where scents are identified (Figure 28-20).
As with lizards, the snake’s body is entirely covered with a tough, impervious skin. The hard scales are set together, sometimes overlapping like shingles on a roof, with the skin folded inward between the scales. The skin is not elastic, and when required to stretch, as it must after the snake has enjoyed a hefty meal, it does so in a novel manner. The skin folds are pulled out straight, leaving the scales separated like islands on the skin.
Snakes have evolved several solutions to the obvious problem of movement without legs. The most typical pattern of movement is lateral undulation (Figure 28-21A). Movement follows an S-shaped path, with the snake propelling itself by exerting lateral force against surface irregularities. The snake seems to “flow,” since the moving loops appear stationary with respect to the ground. Lateral undulatory movement is fast and efficient under most but not all circumstances. Concertina movement (Figure 28-21B) enables a snake to move in a narrow passage, as when climbing a tree by using irregular channels in the bark. The snake extends forward while bracing S-shaped loops against the sides of the channel. To advance in a straight line as when stalking prey, many heavy-bodied snakes employ rectilinear movement. Two or three sections of the body rest on the ground to support the snake’s weight. Intervening sections are lifted free of the ground and pulled forward by muscles (shown in red in Figure 28-21C) that originate on ribs and insert on the ventral skin. Rectilinear movement is a slow but effective way of moving inconspicuously toward prey, even when there are no surface irregularities. Side-winding is a fourth form of movement that enables desert vipers to move with surprising speed across loose, sandy surfaces with minimum surface contact. The sidewinder rattlesnake moves by throwing its body forward in loops with its body lying at an angle of about 60 degrees to its direction of travel.
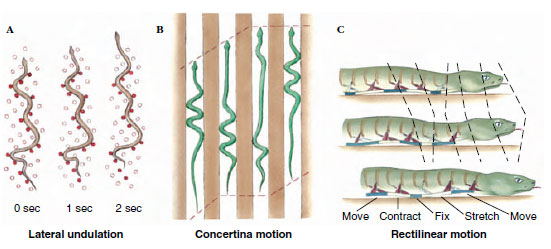 |
| Figure 28-21 Snake locomotion. A, Lateral undulation. B, Concertina motion. C, Rectilinear motion. Refer to text for explanation. |
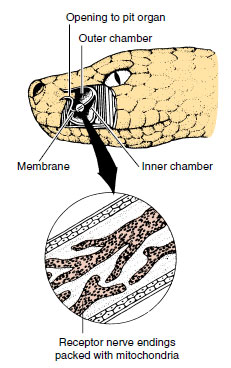 |
| Figure 28-22 Pit organ of rattlesnake, a pit viper. Cutaway shows location of a deep membrane that divides the pit into inner and outer chambers. Heat- sensitive nerve endings are concentrated in the membrane. |
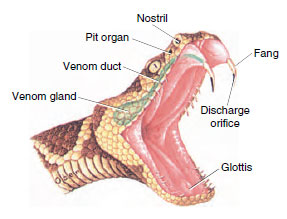 |
| Figure 28-23 Head of rattlesnake showing the venom apparatus. The venom gland, a modified salivary gland, is connected by a duct to the hollow fang. |
The tropical and subtropical countries are the homes of most species of snakes, both of the venomous and nonvenomous varieties. Even in these countries, less than one-third of snakes are venomous. Nonvenomous snakes kill their prey by constriction (Figure 28-24) or by biting and swallowing. Their diet tends to be restricted, many feeding principally on rodents, whereas others feed on fishes, frogs, and insects. Some African, Indian, and neotropical snakes are egg eaters.
 |
| Figure 28-24 Nonvenomous African house snake, Boaedon fuluginosus, constricting a mouse before swallowing it. |
 |
| Figure 28-25 Spectacled, or Indian, cobra, Naja naja. Cobras erect the front of the body and flatten the neck as a threat display and before attacking. Although the cobra’s strike range is limited, all cobras are dangerous because of the extreme toxicity of the venom. |
Even the saliva of harmless snakes possesses limited toxic qualities, and it is logical that there was a natural selection for this toxic tendency as snakes evolved. Snake venoms have traditionally been divided into two types. The neurotoxic type acts mainly on the nervous system, affecting the optic nerves (causing blindness) or the phrenic nerve of the diaphragm (causing paralysis of respiration). The hemorrhagin type breaks down red blood corpuscles and blood vessels and produces extensive hemorrhaging of blood into tissue spaces. In fact, most snake venoms are complex mixtures of various fractions that attack different organs in specific ways; they seldom can be assigned categorically to one or the other of the traditional types.
The toxicity of a venom is measured by the median lethal dose on laboratory animals (LD50). By this standard the venoms of the Australian tiger snake and some of the sea snakes appear to be the most deadly of venomous drop for drop. However, several larger snakes are more dangerous. The aggressive king cobra, which may exceed 5.5 m in length, is the largest and probably the most dangerous of all poisonous snakes. In India and Burma, where snakes come in frequent contact with poorly shod people, some 200,000 snake bites cause more than 10,000 deaths each year. Russell’s viper is responsible for some 1000 deaths each year in Burma alone. The world total for deaths from snakebite is estimated at 25,000 each year.
Most snakes are oviparous (L. ovum, egg, + parere, to bring forth) species that lay their shelled, elliptical eggs beneath rotten logs, under rocks, or in holes in the ground. Most of the remainder, including all the American pit vipers, except the tropical bushmaster, are ovoviviparous (L. ovum, egg, + vivus, living, + parere, to bring forth), giving birth to well-formed young. Very few snakes are viviparous (L. vivus, living, + parere, to bring forth); in these snakes a primitive placenta forms, permitting exchange of materials between the embryonic and maternal bloodstreams. Snakes are able to store sperm and can lay several clutches of fertile eggs at long intervals after a single mating.
 |
| Figure 28-26 Tuatara, Sphenodon sp., a living representative of the order Sphenodonta. This “living fossil” reptile has, on top of the head, a well-developed parietal “eye” with retina, lens, and nervous connections to the brain. Although covered with scales, this third eye is sensitive to light. The parietal eye may have been an important sense organ in early reptiles. The tuatara is found today only on certain islands off the coastline of New Zealand. |
Order Sphenodonta: The Tuatara
The order Sphenodonta is represented by two living species of the genus Sphenodon (Gr. sphenos, wedge, + odontos, tooth) of New Zealand (Figure 28-26). The tuatara is the sole survivor of the sphenodontid lineage that radiated modestly during the early Mesozoic era but declined toward the end of the Mesozoic. Several species of the tuatara were once widespread throughout the two main islands of New Zealand but the two living species are now restricted to small islets of Cook Strait and off the northeast coast of North Island. On some of these islands, under protection by the New Zealand government, they are prospering.
The tuatara is a lizardlike form 66 cm long or less that lives in burrows often shared with petrels. They are slow-growing animals with a long life; one is recorded to have lived 77 years.
The tuatara has captured the interest of zoologists because of numerous features that are almost identical to those of Mesozoic fossils 200 million years ago. These features include a diapsid skull with two temporal openings bounded by complete arches. Tuataras also bear a welldeveloped median parietal eye complete with elements of cornea, lens, and retina (although since it is buried beneath opaque skin this “third eye” can register only changes in light intensity, and its function, if any, remains unknown). In many other respects Sphenodon resembles lizards of the early Mesozoic. Sphenodon represents one of the slowest rates of evolution known among vertebrates.
Order Crocodilia: Crocodiles and Alligators
The modern crocodilians are the only surviving reptiles of the archosaurian lineage that gave rise to the great Mesozoic radiation of dinosaurs and their kin and to the birds. Although modern crocodiles belong to a lineage that began its radiation in the late Cretaceous period, they differ little in structural details from primitive crocodilians of the early Mesozoic. Having remained mostly unchanged for nearly 200 million years, crocodilians face an uncertain future in a world dominated by humans. Modern crocodilians are divided into three families: alligators and caimans, mostly a New World group; crocodiles, which are widely distributed and include the saltwater crocodile, one of the largest living reptiles; and gavials, represented by a single species in India and Burma.
All crocodilians have an elongate, robust, well-reinforced skull and massive jaw musculature arranged to provide a wide gape and rapid, powerful closure. Teeth are set in sockets, a type of dentition called thecodont that was typical of all archosaurs as well as the earliest birds. Another adaptation, found in no other vertebrate except mammals, is a complete secondary palate. This innovation allows crocodilians to breathe when the mouth is filled with water or food (or both). Crocodilians also share a four-chambered heart with birds and mammals.
The estuarine crocodile (Crocodylus porosus), found in southern Asia, and the Nile crocodile (C. niloticus; Figure 28-27A) grow to great size (adults weighing 1000 kg have been reported) and are swift and aggressive. Crocodiles are known to attack animals as large as cattle, deer, and people. Alligators (Figure 28-27B) are usually less aggressive than crocodiles and far less dangerous to humans. Large alligators are powerful animals nevertheless, and adults have almost no enemies but humans. The chink in their formidable armor is the developmental stages. Nests left unguarded by the mother are almost certain to be discovered and raided by any of several mammals that relish eggs, and the young hatchlings may be devoured by large fish.
Alligators are able to make definite vocalizations. The male alligator can give loud bellows in the mating season. In the United States, Alligator mississipiensis (Figure 28-27B) is the only species of alligator; Crocodylus acutus, restricted to extreme southern Florida, is the only species of crocodile.
 |
 |
|
| Figure 28-27 Crocodilians. A, Nile crocodile (Crocodylus niloticus) basking. The fourth tooth of the lower jaw fits outside the slender upper jaw; alligators lack this feature. B, American alligator (Alligator mississipiensis), an increasingly noticeable resident of rivers, bayous, and swamps of the southeastern United States. |
||
Alligators and crocodiles are oviparous. Usually 20 to 50 eggs are laid in a mass of dead vegetation and guarded by the mother. The mother hears vocalizations from the hatching young and responds by opening the nest to allow the hatchlings to escape. As with many turtles and some lizards, incubation temperature of the eggs determines sex ratio of the offspring. However, unlike turtles (low nest temperatures produce only females, whereas high nest temperatures produce only males. This results in highly unbalanced sex ratios in some areas. For example, in one study area in Louisiana, female hatchlings outnumbered males five to one.
Crocodiles and alligators can be distinguished on the basis of head morphology. Crocodiles have a relatively narrow snout, and when their mouths are closed, the fourth lower jaw tooth is visible. Alligators generally have a broader snout, and their fourth lower jaw tooth is hidden by fitting into a notch in the upper jaw (Figure 28-27). Gavials have very narrow snouts, and are largely fish eaters.
Support our developers




