Plastid
Plastids are major organelles found in plants and algae. Plastids are the site of manufacture and storage of important chemical compounds used by the cell. Plastids often contain pigments used in photosynthesis, and the types of pigments present can change or determine the cell's color.Plastids in plants
Plastids are responsible for photosynthesis, storage of products like starch and for the synthesis of many classes of molecules such as fatty acids and terpenes which are needed as cellular building blocks and/or for the function of the plant. Depending on their morphology and function, plastids have the ability to differentiate, or redifferentiate, between these and other forms.
Plant cells with visible chloroplasts.
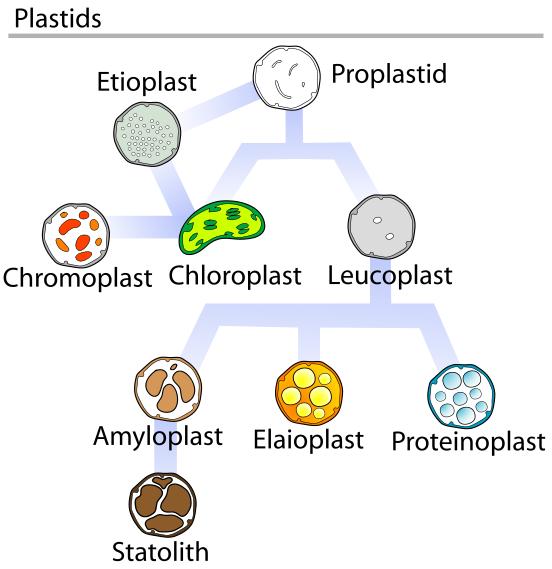
In plants, plastids may differentiate into several forms, depending upon which function they need to play in the cell. Undifferentiated plastids (proplastids) may develop into any of the following plastids:
» Chloroplasts: for photosynthesis; see also etioplasts, the predecessors of chloroplasts
» Chromoplasts: for pigment synthesis and storage
» Leucoplasts: for monoterpene synthesis; leucoplasts sometimes differentiate into more specialized plastids:
» Amyloplasts: for starch storage
» Statoliths: for detecting gravity
» Elaioplasts: for storing fat
» Proteinoplasts: for storing and modifying protein
Each plastid creates multiple copies of the circular 75-250 kilo bases plastid genome. The number of genome copies per plastid is flexible, ranging from more than 1000 in rapidly dividing cells, which generally contain few plastids, to 100 or fewer in mature cells, where plastid divisions has given rise to a large number of plastids.
Plastid DNA exists as large protein-DNA complexes associated with the inner envelope membrane and called 'plastid nucleoids'. Each nucleoid particle may contain more than 10 copies of the plastid DNA. The proplastid contains a single nucleoid located in the centre of the plastid. The developing plastid has many nucleoids, localized at the periphery of the plastid, bound to the inner envelope membrane. During the development of proplastids to chloroplasts, and when plastids convert from one type to another, nucleoids change in morphology, size and location within the organelle. The remodelling of nucleoids is believed to occur by modifications to the composition and abundance of nucleoid proteins.
In plant cells long thin protuberances called stromules sometimes form and extend from the main plastid body into the cytosol and interconnect several plastids. Proteins, and presumably smaller molecules, can move within stromules. Most cultured cells that are relatively large compared to other plant cells have very long and abundant stromules that extend to the cell periphery.
Plastids in algae
In algae, the term leucoplast (leukoplast) is used for all unpigmented plastids. Their function differ from the leukoplasts in plants. Etioplast, amyloplast and chromoplast are plant-specific and do not occur in algae. Algal plastids may also differ from plant plastids in that they contain pyrenoids.Inheritance of plastids
Most plants inherit the plastids from only one parent. Angiosperms generally inherit plastids from the mother, while many gymnosperms inherit plastids from the father. Algae also inherit plastids from only one parent. The plastid DNA of the other parent is thus completely lost.In normal intraspecific crossings (resulting in normal hybrids of one species), the inheritance of plastid DNA appears to be quite strictly 100% uniparental. In interspecific hybridisations, however, the inheritance of plastids appears to be more erratic. Although plastids inherit mainly maternally in interspecific hybridisations, there are many reports of hybrids of flowering plants that contain plastids of the father.
Origin of plastids
Plastids are thought to have originated from endosymbiotic cyanobacteria. They developed around 1500 mya and allowed eukaryotes to carry out oxygenic photosynthesis. Due to a split-up into three evolutionary lineages, the plastids are named differently: chloroplasts in green algae and plants, rhodoplasts in red algae and cyanelles in the glaucophytes. The plastids differ by their pigmentation, but also in ultrastructure. The chloroplasts e.g. have lost all phycobilisomes, the light harvesting complexes found in cyanobacteria, red algae and glaucophytes, but - only in plants and in closely related green algae - contain stroma and grana thylakoids. The glaucocystophycean plastid - in contrast to the chloroplasts and the rhodoplasts - is still surrounded by the remains of the cyanobacterial cell wall. All these primary plastids are surrounded by two membranes.Some dinoflagellates take up algae as food and keep the plastid of the digested alga to profit from the photosynthesis; after a while the plastids are also digested. These captured plastids are known as kleptoplastids.
Sources
- A Novel View of Chloroplast Structure: contains fluorescence images of chloroplasts and stromules as well as an easy to read chapter.- Continuous expression in tobacco leaves of a Brassica napus PEND homologue blocks differentiation of plastids and development of palisade cells Wycliffe et al., 2005. The Plant Journal Volume 44 Issue 1 Page 1.
- Birky, C. W. 2001. The inheritance of genes in mitochondria and chloroplasts: laws, mechanisms and models. Annual Review of Genetics 35: 125-148. URL
References
- Hedges, S. Blair et al. (2004) "A molecular timescale of eukaryote evolution and the rise of complex multicellular life" BMC Evolutionary Biology 4:2Further reading
- Bhattacharya, D. (Ed.) 1997 Origins of Algae and their Plastids. Springer-Verlag/Wein, New York.Support our developers




