Plant Hormone
Contents
» Characteristics» Classes of plant hormones
» Abscisic acid
» Auxins
» Cytokinins
» Ethylene
» Gibberellins
» Other known hormones
» Potential medical applications
» Hormones and plant propagation
» Seed dormancy
» References
Characteristics
The word hormone is derived from Greek and means 'set in motion.'
Lacking the plant hormone auxin, causes abnormal growth (right)
Plant hormones are not nutrients, but chemicals that in small amounts promote and influence the growth, development, and differentiation of cells and tissues. The biosynthesis of plant hormones within plant tissues is often diffuse and not always localized. Plants lack glands to produce and store hormones, because, unlike animals, which have two circulatory systems (lymphatic and cardiovascular) powered by a heart that moves fluids around the body, plants use more passive means to move chemicals around the plant. Plants utilize simple chemicals as hormones, which move more easily through the plant's tissues. They are often produced and used on a local basis within the plant body, plant cells even produce hormones that affect different regions of the cell producing the hormone.
Not all plant cells respond to hormones, but those cells that do, are programmed to respond at specific points in their growth cycle. The greatest effects occur at specific stages during the cell's life, with diminished effects occurring before or after this period. Plants need hormones at very specific times during plant growth and at specific locations. They also need to disengage the effects that hormones have when they are no longer needed. The production of hormones occurs very often at sites of active growth within the meristems, before cells have fully differentiated. After production they are sometimes moved to other parts of the plant where they cause an immediate effect or they can be stored in cells to be released later. Plants use different pathways to regulate internal hormone quantities and moderate their effects; they can regulate the amount of chemicals used to biosynthesize hormones. They can store them in cells, inactivate them, or cannibalise already-formed hormones by conjugating them with carbohydrates, amino acids or peptides. Plants can also break down hormones chemically, effectively destroying them. Plants also move hormones around the plant diluting their concentrations.
Classes of plant hormones
It is generally accepted that there are five major classes of plant hormones, some of which are made up of many different chemicals that can vary in structure from one plant to the next. The chemicals are each grouped together into one of these classes based on their structural similarities and on their effects on plant physiology. Other plant hormones and growth regulators are not easily grouped into these classes, they exist naturally or are synthesized by humans or other organisms, including chemicals that inhibit plant growth or interrupt the physiological processes within plants. Each class has positive as well as inhibitory functions, and most often work in tandem with each other, with varying ratios of one or more interplaying to affect growth regulation.The five major classes are:
Abscisic acid
Abscisic acid also called ABA, was discovered and researched under two different names before its chemical properties were fully known, it was called dormin and abscicin II. Once it was determined that the two latter named compounds were the same, it was named abscisic acid. The name "abscisic acid" was given because it was found in high concentrations in newly-abscissed or freshly-fallen leaves.This class of PGR is composed of one chemical compound normally produced in the leaves of plants, originating from chloroplasts, especially when plants are under stress. In general, it acts as an inhibitory chemical compound that affects bud growth, seed and bud dormancy. It mediates changes within the apical meristem causing bud dormancy and the alteration of the last set of leaves into protective bud covers. Since it was found in freshly-abscissed leaves, it was thought to play a role in the processes of natural leaf drop but further research has disproven this. In plant species from temperate parts of the world it plays a role in leaf and seed dormancy by inhibiting growth, but, as it is dissipated from seeds or buds, growth begins. In other plants, as ABA levels decrease, growth then commences as gibberellin levels increase. Without ABA, buds and seeds would start to grow during warm periods in winter and be killed when it froze again. Since ABA dissipates slowly from the tissues and its effects take time to be offset by other plant hormones, there is a delay in physiological pathways that provide some protection from premature growth. It accumulates within seeds during fruit maturation, preventing seed germination within the fruit, or seed germination before winter. Abscisic acid's effects are degraded within plant tissues during cold temperatures or by its removal by water washing in out of the tissues, releasing the seeds and buds from dormancy.
Auxins
Auxins are compounds that positively influence cell enlargement, bud formation and root initiation. They also promote the production of other hormones and in conjunction with cytokinins, they control the growth of stems, roots, flowers and fruits. Auxins were the first class of growth regulators discovered. They affect cell elongation by altering cell wall plasticity. Auxins decrease in light and increase where its dark. They stimulate cambium cells to divide and in stems cause secondary xylem to differentiate. Auxins act to inhibit the growth of buds lower down the stems (apical dominance), and also to promote lateral and adventitious root development and growth. Auxins promote flower initiation, converting stems into flowers. Leaf abscission is initiated by the growing point of a plant ceasing to produce auxins. Auxins in seeds regulate specific protein synthesis, as they develop within the flower after pollination, causing the flower to develop a fruit to contain the developing seeds. Auxins are toxic to plants in large concentrations; they are most toxic to dicots and less so to monocots. Because of this property, synthetic auxin herbicides including 2,4-D and 2,4,5-T have been developed and used for weed control. Auxins, especially 1-Naphthaleneacetic acid (NAA) and Indole-3-butyric acid (IBA), are also commonly applied to stimulate root growth when taking cuttings of plants. The most common auxin found in plants is indoleacetic acid or IAA.Cytokinins
Cytokinins or CKs are a group of chemicals that influence cell division and shoot formation. They were called kinins in the past when the first cytokinins were isolated from yeast cells. They also help delay senescence or the aging of tissues, are responsible for mediating auxin transport throughout the plant, and affect internodal length and leaf growth. They have a highly-synergistic effect in concert with auxins and the ratios of these two groups of plant hormones affect most major growth periods during a plant's lifetime. Cytokinins counter the apical dominance induced by auxins; they in conjunction with ethylene promote abscission of leaves, flower parts and fruits.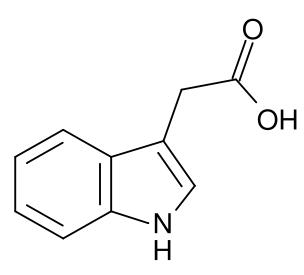
The auxin indoleacetic acid
Ethylene
Ethylene is a gas that forms from the breakdown of methionine, which is in all cells. Ethylene has very limited solubility in water and does not accumulate within the cell but diffuses out of the cell and escapes out of the plant. Its effectiveness as a plant hormone is dependent on its rate of production versus its rate of escaping into the atmosphere. Ethylene is produced at a faster rate in rapidly growing and dividing cells, especially in darkness.New growth and newly-germinated seedlings produce more ethylene than can escape the plant, which leads to elevated amounts of ethylene, inhibiting leaf expansion. As the new shoot is exposed to light, reactions by phytochrome in the plant's cells produce a signal for ethylene production to decrease, allowing leaf expansion. Ethylene affects cell growth and cell shape; when a growing shoot hits an obstacle while underground, ethylene production greatly increases, preventing cell elongation and causing the stem to swell. The resulting thicker stem can exert more pressure against the object impeding its path to the surface. If the shoot does not reach the surface and the ethylene stimulus becomes prolonged, it affects the stems natural geotropic response, which is to grow upright, allowing it to grow around an object. Studies seem to indicate that ethylene affects stem diameter and height: When stems of trees are subjected to wind, causing lateral stress, greater ethylene production occurs, resulting in thicker, more sturdy tree trunks and branches. Ethylene affects fruit-ripening: Normally, when the seeds are mature, ethylene production increases and builds-up within the fruit, resulting in a climacteric event just before seed dispersal. The nuclear protein ETHYLENE INSENSITIVE2 (EIN2) is regulated by ethylene production, and, in turn, regulates other hormones including ABA and stress hormones.
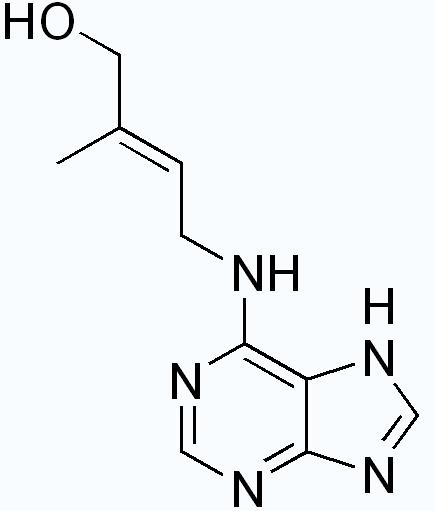
The cytokinin zeatin , Zea, in which it was first discovered in immature kernels.
Gibberellins
Gibberellins or GAs include a large range of chemicals that are produced naturally within plants and by fungi. They were first discovered when Japanese researchers, including Eiichi Kurosawa, noticed a chemical produced by a fungus called Gibberella fujikuroi that produced abnormal growth in rice plants. Gibberellins are important in seed germination, affecting enzyme production which mobilizes food production used for growth of new cells. This is done by modulating chromosomal transcription. In seedlings a layer of cells called the aleurone layer wraps around the endosperm tissue: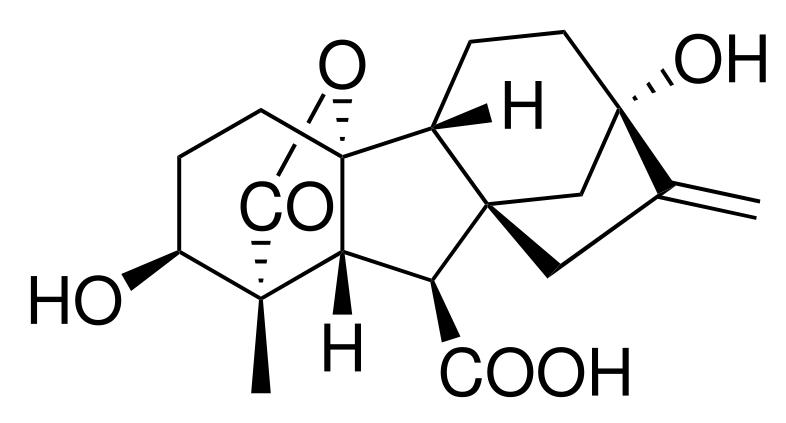
Gibberellin A1
Other known hormones
Other identified plant growth regulators include:» Brassinolides - plant steroids that are chemically similar to animal steroid hormones. First isolated from pollen of the mustard family and extensively studied in Arabidopsis. They promote cell elongation and cell division, differentiation of xylem tissues, and inhibit leaf abscission. Plants that are deficient in brassinolides suffer from dwarfism.
» Salicylic acid - activates genes in some plants that produce chemicals that aid in the defense against pathogenic invaders.
» Jasmonates - are produced from fatty acids and seem to promote the production of defense proteins that are used to fend off invading organisms. They are believed to also have a role in seed germination, and affect the storage of protein in seeds, and seem to affect root growth.
» Plant peptide hormones - encompass all small secreted peptides that are involved in cell-to-cell signaling. These small peptide hormones play crucial roles in plant growth and development, including defense mechanisms, the control of cell division and expansion, and pollen self-incompatibility.
» Polyamines - are strongly basic molecules with low molecular weight that have been found in all organisms studied thus far. They are essential for plant growth and development and affect the process of mitosis and meiosis.
» Nitric oxide (NO) - serves as signal in hormonal and defense responses.
» Strigolactones, implicated in the inhibition of shoot branching.
Potential medical applications
Plant stress hormones activate cellular responses, including cell death, to diverse stress situations in plants. Researchers have found that some plant stress hormones share the ability to adversely affect human cancer cells. For example, sodium salicylate has been found to suppress proliferation of lymphoblastic leukemia, prostate, breast, and melanoma human cancer cells. Jasmonic acid, a plant stress hormone that belongs to the jasmonate family, induced death in lymphoblastic leukemia cells. Methyl jasmonate has been found to induce cell death in a number of cancer cell lines.Hormones and plant propagation
Synthetic plant hormones or PGRs are commonly used in a number of different techniques involving plant propagation from cuttings, grafting, micropropagation, and tissue culture.The propagation of plants by cuttings of fully-developed leaves, stems, or roots is performed by gardeners utilizing auxin as a rooting compound applied to the cut surface; the auxins are taken into the plant and promote root initiation. In grafting, auxin promotes callus tissue formation, which joins the surfaces of the graft together. In micropropagation, different PGRs are used to promote multiplication and then rooting of new plantlets. In the tissue-culturing of plant cells, PGRs are used to produce callus growth, multiplication, and rooting.
Seed dormancy
Plant hormones affect seed germination and dormancy by affecting different parts of the seed.Embryo dormancy is characterized by a high ABA/GA ratio, whereas the seed has a high ABA sensitivity and low GA sensitivity. To release the seed from this type of dormancy and initiate seed germination, an alteration in hormone biosynthesis and degradation towards a low ABA/GA ratio, along with a decrease in ABA sensitivity and an increase in GA sensitivity needs to occur.
ABA controls embryo dormancy, and GA embryo germination. Seed coat dormancy involves the mechanical restriction of the seed coat, this along with a low embryo growth potential, effectively produces seed dormancy. GA releases this dormancy by increasing the embryo growth potential, and/or weakening the seed coat so the radical of the seedling can break through the seed coat. Different types of seed coats can be made up of living or dead cells and both types can be influenced by hormones; those composed of living cells are acted upon after seed formation while the seed coats composed of dead cells can be influenced by hormones during the formation of the seed coat. ABA affects testa or seed coat growth characteristics, including thickness, and effects the GA-mediated embryo growth potential. These conditions and effects occur during the formation of the seed, often in response to environmental conditions. Hormones also mediate endosperm dormancy: Endosperm in most seeds is composed of living tissue that can actively respond to hormones generated by the embryo. The endosperm often acts as a barrier to seed germination, playing a part in seed coat dormancy or in the germination process. Living cells respond to and also affect the ABA/GA ratio, and mediate cellular sensitivity; GA thus increases the embryo growth potential and can promote endosperm weakening. GA also affects both ABA-independent and ABA-inhibiting processes within the endosperm.
References
- Srivastava, L. M. 2002. Plant growth and development: hormones and environment. Amsterdam: Academic Press. Page 140.- Helgi Öpik, Stephen A. Rolfe The Physiology of Flowering Plants Published 2005 Cambridge University Press Plant physiology Page 191
- Srivastava, L. M. 2002. Plant growth and development hormones and environment. Amsterdam: Academic Press. Page 143.
- Rost, Thomas L., and T. Elliot Weier. 1979. Botany: a brief introduction to plant biology. New York: Wiley. Pages 155-170.
- Dormancy termination of western white pine ( Pinus monticola Dougl. Ex D. Don) seeds is associated with changes in abscisic acid metabolism Journal Planta Publisher Springer Berlin / Heidelberg
- Dynamic analysis of ABA accumulation in relation to the rate of ABA catabolism in maize tissues under water deficit. Huibo Ren1 *, Zhihui Gao1 *, Lin Chen1 *, Kaifa Wei1, Jing Liu1, Yijuan Fan1, William J. Davies2, Wensuo Jia1,{dagger} and Jianhua Zhang3,{dagger} JXB Advance Access originally published online on September 18, 2006. Journal of Experimental Botany 2007 58(2):211-219;
- Decreased root hydraulic conductivity reduces leaf water potential, initiates stomatal closure and slows leaf expansion in flooded plants of castor oil (Ricinus communis) despite diminished delivery of ABA from the roots to shoots in xylem sap. Else M.A.1; Coupland D.2; Dutton L.3; Jackson M.B.3 Physiologia Plantarum, Volume 111, Number 1, January 2001 , pp. 46-54(9) Blackwell Publishing
- Reactive oxygen species and nitric oxide are involved in ABA inhibition of stomatal opening. YAN, JIUPIANG; TSUICHIHARA, NOBUE; ETOH, TAKEOMI; IWAI, SUMIO Plant, Cell & Environment, Volume 30, Number 10, October 2007 , pp. 1320-1325(6) Blackwell Publishing
- Role of Abscisic Acid in Seed Dormancy Journal Journal of Plant Growth Regulation Publisher Springer New York ISSN 0721-7595 (Print) 1435-8107 (Online) Issue Volume 24, Number 4 / December, 2005 Category Thematic Article
- Classification of auxin related compounds based on similarity of their interaction fields: Extension to a new set of compounds Tomic, S.1,2, Gabdoulline, R.R.1, Kojic-Prodic, B.2 and Wade, R.C.11 - European Molecular Biology Laboratory, 69012 Heidelberg, Germany 2Institute Rudjer Boskovic, HR-10000 Zagreb, Croatia
- A gene encoding a protein modified by the phytohormone indoleacetic acid. Alexander Walz, Seijin Park, Janet P. Slovin, Jutta Ludwig-Müller, Yoshie S. Momonoki, and Jerry D. Cohen Proc Natl Acad Sci U S A. 2002 February 5; 99(3): 1718–1723.
- Deborah L. Sipes and John W. Einset. Cytokinin stimulation of abscission in lemon pistil explants Journal Journal of Plant Growth Regulation Publisher Springer New York.
- Arabidopsis EIN2 modulates stress response through abscisic acid response pathway. Authors: Wang, Youning; Liu, Chuang; Li, Kexue; Sun, Feifei; Hu, Haizhou; Li, Xia1; Zhao, Yankun; Han, Chunyu; Zhang, Wensheng; Duan, Yunfeng; Liu, Mengyu. Source: Plant Molecular Biology, Volume 64, Number 6, August 2007 , pp. 633-644(12) Springer
- Grennan, Aleel K (2006), "Gibberellin Metabolism Enzymes in Rice", Plant Physiology 141 (2): 524.
- A comparative study of the effects of abscisic acid and methyl jasmonate on seedling growth of rice Authors: Tsai F-Y.1; Lin C.C.1; Kao C.H.1 Plant Growth Regulation, Volume 21, Number 1, January 1997 , pp. 37-42(6) Springer
- Lindsey, Keith; Casson, Stuart; Chilley, Paul. (2002), "Peptides:new signalling molecules in plants", Trends in Plant Science 7 (2): 78-83.
- Gomez-roldan, Victoria; Fermas, Soraya; Brewer, Philip B.; Puech-pag (2008), "Strigolactone inhibition of shoot branching", Nature 455 (7210): 189.
Support our developers




