Cyanobacteria
Cyanobacteria, also known as blue-green algae, blue-green bacteria or Cyanophyta, is a phylum of bacteria that obtain their energy through photosynthesis. The name "cyanobacteria" comes from the color of the bacteria (Greek: κυανός (kyanós) = blue). They are a significant component of the marine nitrogen cycle and an important primary producer in many areas of the ocean, but are also found in habitats other than the marine environment; in particular cyanobacteria are known to occur in both freshwater and hypersaline inland lakes.Stromatolites of fossilized oxygen-producing cyanobacteria have been found from 3.5 to 2.8 billion years ago. The ability of cyanobacteria to perform oxygenic photosynthesis is thought to have converted the early reducing atmosphere into an oxidizing one, which dramatically changed the composition of life forms on Earth by provoking an explosion of biodiversity and leading to the near-extinction of oxygen-intolerant organisms. Chloroplasts in plants and eukaryotic algae have evolved from cyanobacteria via endosymbiosis.
Contents
⇒ Forms
⇒ Photosynthesis
⇒ Relationship to chloroplasts
⇒ Relationship to Earth history
⇒ Classification
⇒ Biotechnology and applications
⇒ Health risks
⇒ See also
⇒ References
⇒ Further reading
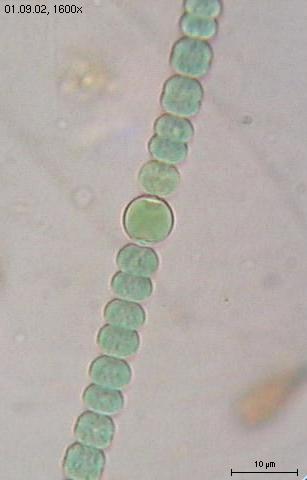 |
|
Cyanobacteria: Anabaena sphaerica |
|
Forms
Cyanobacteria are found in almost every conceivable environment, from oceans to fresh water to bare rock to soil. Most are found in fresh water, while others are marine, occur in damp soil, or even temporarily moistened rocks in deserts. A few are endosymbionts in lichens, plants, various protists, or sponges and provide energy for the host. Some live in the fur of sloths, providing a form of camouflage.
Cyanobacteria include unicellular and colonial species. Colonies may form filaments, sheets or even hollow balls. Some filamentous colonies show the ability to differentiate into several different cell types: vegetative cells, the normal, photosynthetic cells that are formed under favorable growing conditions; akinetes, the climate-resistant spores that may form when environmental conditions become harsh; and thick-walled heterocysts, which contain the enzyme nitrogenase, vital for nitrogen fixation.
 |
|
Colonies of Nostoc pruniforme |
|
Heterocysts may also form under the appropriate environmental conditions (anoxic) wherever nitrogen is necessary. Heterocyst-forming species are specialized for nitrogen fixation and are able to fix nitrogen gas, which cannot be used by plants, into ammonia (NH3), nitrites (NO3−) or nitrates (NO3−), which can be absorbed by plants and converted to protein and nucleic acids. The rice paddies of Asia, which produce about 75% of the world's rice, could not do so were it not for healthy populations of nitrogen-fixing cyanobacteria in the rice paddy fertilizer.
Many cyanobacteria also form motile filaments, called hormogonia, that travel away from the main biomass to bud and form new colonies elsewhere. The cells in a hormogonium are often thinner than in the vegetative state, and the cells on either end of the motile chain may be tapered. In order to break away from the parent colony, a hormogonium often must tear apart a weaker cell in a filament, called a necridium.
Each individual cell of a cyanobacterium typically has a thick, gelatinous cell wall. They differ from other gram-negative bacteria in that the quorum sensing molecules autoinducer-2 and acyl-homoserine lactones are absent. They lack flagella, but hormogonia and some unicellular species may move about by gliding along surfaces. In water columns some cyanobacteria float by forming gas vesicles, like in archaea.
Some of these organisms contribute significantly to global ecology and the oxygen cycle. The tiny marine cyanobacterium Prochlorococcus was discovered in 1986 and accounts for more than half of the photosynthesis of the open ocean.
Photosynthesis
Cyanobacteria have an elaborate and highly organized system of internal membranes which function in photosynthesis. Photosynthesis in cyanobacteria generally uses water as an electron donor and produces oxygen as a by-product, though some may also use hydrogen sulfide as occurs among other photosynthetic bacteria. Carbon dioxide is reduced to form carbohydrates via the Calvin cycle. In most forms the photosynthetic machinery is embedded into folds of the cell membrane, called thylakoids. The large amounts of oxygen in the atmosphere are considered to have been first created by the activities of ancient cyanobacteria. Due to their ability to fix nitrogen in aerobic conditions they are often found as symbionts with a number of other groups of organisms such as fungi (lichens), corals, pteridophytes (Azolla), angiosperms (Gunnera) etc.
Cyanobacteria are the only group of organisms that are able to reduce nitrogen and carbon in aerobic conditions, a fact that may be responsible for their evolutionary and ecological success. The water-oxidizing photosynthesis is accomplished by coupling the activity of photosystem (PS) II and I (Z-scheme). In anaerobic conditions, they are also able to use only PS I — cyclic photophosphorylation — with electron donors other than water (hydrogen sulfide, thiosulphate, or even molecular hydrogen) just like purple photosynthetic bacteria. Furthermore, they share an archaeal property, the ability to reduce elemental sulfur by anaerobic respiration in the dark. Their photosynthetic electron transport shares the same compartment as the components of respiratory electron transport. Actually, their plasma membrane contains only components of the respiratory chain, while the thylakoid membrane hosts both respiratory and photosynthetic electron transport.
Attached to thylakoid membrane, phycobilisomes act as light harvesting antennae for the photosystems . The phycobilisome components (phycobiliproteins) are responsible for the blue-green pigmentation of most cyanobacteria. The variations to this theme is mainly due to carotenoids and phycoerythrins which give the cells the red-brownish coloration. In some cyanobacteria, the color of light influences the composition of phycobilisomes. In green light, the cells accumulate more phycoerythrin, whereas in red light they produce more phycocyanin. Thus the bacteria appear green in red light and red in green light. This process is known as complementary chromatic adaptation and is a way for the cells to maximize the use of available light for photosynthesis.
A few genera, however, lack phycobilisomes and have chlorophyll b instead (Prochloron, Prochlorococcus, Prochlorothrix). These were originally grouped together as the prochlorophytes or chloroxybacteria, but appear to have developed in several different lines of cyanobacteria. For this reason they are now considered as part of the cyanobacterial group.
Relationship to chloroplasts
Chloroplasts found in eukaryotes (algae and plants) likely evolved from an endosymbiotic relation with cyanobacteria. This endosymbiotic theory is supported by various structural and genetic similarities. Primary chloroplasts are found among the green plants, where they contain chlorophyll b, and among the red algae and glaucophytes, where they contain phycobilins. It now appears that these chloroplasts probably had a single origin, in an ancestor of the clade called Primoplantae. Other algae likely took their chloroplasts from these forms by secondary endosymbiosis or ingestion.
It was once thought that the mitochondria in eukaryotes also developed from an endosymbiotic relationship with cyanobacteria; however, it is now suspected that this evolutionary event occurred when aerobic bacteria were engulfed by anaerobic host cells. Mitochondria are believed to have originated not from cyanobacteria but from an ancestor of Rickettsia.
Relationship to Earth History
 |
The biochemical capacity to use water as the source for electrons in photosynthesis evolved once, in a common ancestor of extant cyanobacteria. The geologic record indicates that this transforming event took place early in our planet's history, at least 2450-2320 million years ago (Ma), and probably much earlier. Geobiological interpretation of Archean (>2500 Ma) sedimentary rocks remains a challenge; available evidence indicates that life existed 3500 Ma, but the question of when oxygenic photosynthesis evolved continues to engender debate and research. A clear paleontological window on cyanobacterial evolution opened about 2000 Ma, revealing an already diverse biota of blue-greens. Cyanobacteria remained principal primary producers throughout the Proterozoic Eon (2500-543 Ma), in part because the redox structure of the oceans favored photautotrophs capable of nitrogen fixation. Green algae joined blue-greens as major primary producers on continental shelves near the end of the Proterozoic, but only with the Mesozoic (251-65 Ma) radiations of dinoflagellates, coccolithophorids, and diatoms did primary production in marine shelf waters take modern form. |
|
Oncolites; Guilmette Formation (Late Devonian) near Hancock Summit, Pahranagat Range, Nevada. |
||
The most common cyanobacterial structures in the fossil record include stromatolites and oncolites. Cyanobacteria remain critical to marine ecosystems as primary producers in oceanic gyres, as agents of biological nitrogen fixation, and, in modified form, as the plastids of marine algae.
Classification
The cyanobacteria were traditionally classified by morphology into five sections, referred to by the numerals I-V. The first three - Chroococcales, Pleurocapsales, and Oscillatoriales - are not supported by phylogenetic studies. However, the latter two - Nostocales and Stigonematales - are monophyletic, and make up the heterocystous cyanobacteria. The members of Chroococales are unicellular and usually aggregate in colonies. The classic taxonomic criterion has been the cell morphology and the plane of cell division. In Pleurocapsales, the cells have the ability to form internal spores (baeocytes). The rest of the sections include filamentous species. In Oscillatoriales, the cells are uniseriately arranged and do not form specialized cells (akinetes and heterocysts). In Nostocales and Stigonematales the cells have the ability to develop heterocysts in certain conditions. Stigonematales, unlike Nostocales, includes species with truly branched trichomes. Most taxa included in the phylum or division Cyanobacteria have not yet been validly published under the Bacteriological Code. Except:
⇒ The classes Chroobacteria, Hormogoneae and Gloeobacteria
⇒ The orders Chroococcales, Gloeobacterales, Nostocales, Oscillatoriales, Pleurocapsales & Stigonematales
⇒ The families Prochloraceae and Prochlorotrichaceae
⇒ The genera Halospirulina, Planktothricoides, Prochlorococcus, Prochloron, Prochlorothrix.
Biotechnology and applications
The unicellular cyanobacterium Synechocystis sp. PCC6803 was the third prokaryote and first photosynthetic organism whose genome was completely sequenced. It continues to be an important model organism. The smallest genomes have been found in Prochlorococcus spp. (1.7 Mb) and the largest in Nostoc punctiforme (9 Mb). Those of Calothrix spp. are estimated at 12-15 Mb, as large as yeast.
Some cyanobacteria are sold as food, notably Aphanizomenon flos-aquae and Arthrospira platensis (Spirulina).
Health risks
Certain cyanobacteria produce cyanotoxins like anatoxin-a, anatoxin-as, aplysiatoxin, cylindrospermopsin, domoic acid, microcystin LR, nodularin R (from Nodularia), or saxitoxin. Sometimes a mass-reproduction of cyanobacteria results in algal blooms.
These toxins can be neurotoxins, hepatotoxins, cytotoxins, and endotoxins, and can be dangerous to animals and humans. Several cases of human poisoning have been documented but a lack of knowledge prevents an accurate assessment of the risks.
See also
⇒ Archean
⇒ Cyanobiont
⇒ Hypolith
⇒ Oxygen Catastrophe
⇒ Proterozoic
References
| - | Ahoren Oren (2004). "A proposal for further integration of the cyanobacteria under the Bacteriological Code". Int. J. Syst. Evol. Microbiol. 54: 1895–1902. |
| - | Olson JM (2006). "Photosynthesis in the Archean era". Photosyn. Res. 88 (2): 109–17. |
| - | J. Sun, et al. (2004). "Is autoinducer-2 a universal signal for interspecies communication? A comparative genomic and phylogenetic analysis of the synthesis and signal transduction pathways". BMC Evol. Biol. 4: 36. |
| - | E. Dittmann, et al. (2001). "Altered expression of two light-dependent genes in a microcystin-lacking mutant of Microcystis aeruginosa PCC7806". Microbiology 147: 3113–3119. |
| - | Enrique Flores AH (2008). The Cyanobacteria: Molecular Biology, Genomics and Evolution. Horizon. pp. 3. |
| - | Herrero A and Flores E (editor). (2008). The Cyanobacteria: Molecular Biology, Genomics and Evolution (1st ed. ed.). Caister Academic Press. |
| - | T. Kaneko, et al. (1996). "Kaneko, T. et al. (1996) Sequence analysis of the genome of the unicellular cyanobacterium Synechocystis sp. strain PCC6803. II. Sequence determination of the entire genome and assignment of potential protein-coding regions". DNA Res. 3: 109–136. |
| - | Tabei Y, Okada K, Tsuzuki M (2007). "Sll1330 controls the expression of glycolytic genes in Synechocystis sp. PCC 6803". Biochem. Biophys. Res. Commun. 355 (4): 1045–50. |
| - | G. Rocap, et al. (2003). "Genome divergence in two Prochlorococcus ecotypes reflects oceanic niche differentiation". Nature 424: 1042–1047. doi:10.1038/nature01947. |
| - | A. Dufresne, et al. (2003). "Genome sequence of the cyanobacterium Prochlorococcus marinus SS120, a nearly minimal oxyphototrophic genome.". Proc. Natl Acad. Sci. USA 100: 10020–10025. |
| - | J.C. Meeks, et al. (2001). "An overview of the genome of Nostoc punctiforme, a multicellular, symbiotic cyanobacterium". Photosynth. Res. 70: 85–106 A number of important advances have occurred in cyanobacterial biotechnology in the recent years. World wide attention is drawn towards cyanobacteria for their possible use in mariculture, food, feed, fuel, fertilizer, colourant, production of various secondary metabolites including vitamins, toxins, enzymes, pharmaceuticals, pharmacological probes and pollution abatement. Only a few cyanobacterial strains (including Spirulina) have been well-characterized or exploited commercially (Thajuddin and Subramanian. Cyanobacterial biodiversity and potential applications in biotechnology. CURRENT SCIENCE, VOL. 89, NO. 1, 10 JULY 2005). |
| - | M. Herdman, et al. (1979). "Genome size of cyanobacteria". J. Gen. Microbiol. 111: 73–85. |
| - | Spolaore P, Joannis-Cassan C, Duran E, Isambert A (2006). "Commercial applications of microalgae". J. Biosci. Bioeng. 101 (2): 87–96. |
Further Reading
| - | Gillian Cribbs (1997) Nature's Superfood, the Blue-Green Algae Revolution. Newleaf. |
| - | Marshall Savage, (1992, 1994) The Millennial Project: Colonizing the Galaxy in Eight Easy Steps. Little, Brown. |
| - | Fogg, G.E., Stewart, W.D.P., Fay, P.and Walsby, A.E. 1973. The Blue-green Algae. Academic Press, London and New York. |
| - | "Architects of the earth's atmosphere." Introduction to the Cyanobacteria. University of California, Berkeley. 03 Feb. 2006. |
| - | Whitton, B.A. Phylum Cyanophyta (Cyanobacteria). in The Freshwater Algal Flora of the British Isles. Cambridge University Press, Cambridge. |
Support our developers




