Plant Breeding
Plant breeding has been practiced for thousands of years, since near the beginning of human civilization. It is now practiced worldwide by individuals such as gardeners and farmers, or by professional plant breeders employed by organizations such as government institutions, universities, crop-specific industry associations or research centers.
International development agencies believe that breeding new crops is important for ensuring food security by developing new varieties that are higher-yielding, resistant to pests and diseases, drought-resistant or regionally adapted to different environments and growing conditions.
Contents
» Domestication» Classical plant breeding
» Before World War II
» After World War II
» Modern plant breeding
» Steps of Plant Breeding
» Marker assisted selection
» Reverse Breeding and Doubled Haploids (DH)
» Genetic modification
» Issues and concerns
» Participatory Plant Breeding
» List of notable plant breeders
» Notes
» References
Domestication
Plant breeding in certain situations may lead the domestication of wild plants. Domestication of plants is an artificial selection process conducted by humans to produce plants that have more desirable traits than wild plants, and which renders them dependent on artificial (usually enhanced) environments for their continued existence. The practice is estimated to date back 9,000-11,000 years. Many crops in present day cultivation are the result of domestication in ancient times, about 5,000 years ago in the Old World and 3,000 years ago in the New World. In the Neolithic period, domestication took a minimum of 1,000 years and a maximum of 7,000 years. Today, all of our principal food crops come from domesticated varieties. Almost all the domesticated plants used today for food and agriculture were domesticated in the centers of origin. In these centers there is still a great diversity of closely related wild plants, the so called crop wild relatives that can also be used for improving modern cultivars by plant breeding.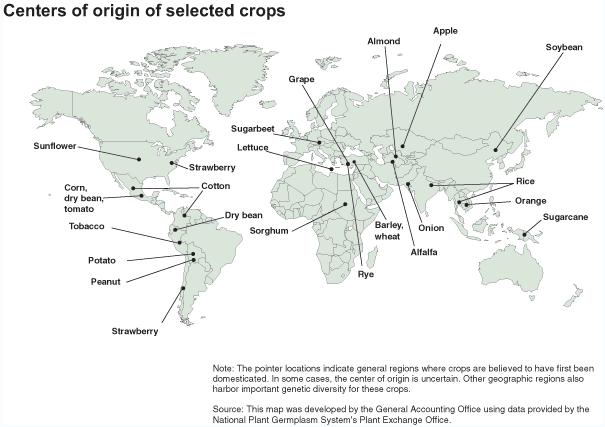
This map shows the sites of domestication for a number of crops. Places where crops were initially domesticated are called centers of origin
Classical plant breeding
Classical plant breeding uses deliberate interbreeding (crossing) of closely or distantly related individuals to produce new crop varieties or lines with desirable properties. Plants are crossbred to introduce traits/genes from one variety or line into a new genetic background. For example, a mildew-resistant pea may be crossed with a high-yielding but susceptible pea, the goal of the cross being to introduce mildew resistance without losing the high-yield characteristics. Progeny from the cross would then be crossed with the high-yielding parent to ensure that the progeny were most like the high-yielding parent, (backcrossing). The progeny from that cross would then be tested for yield and mildew resistance and high-yielding resistant plants would be further developed.Plants may also be crossed with themselves to produce inbred varieties for breeding.
Classical breeding relies largely on homologous recombination between chromosomes to generate genetic diversity. The classical plant breeder may also makes use of a number of in vitro techniques such as protoplast fusion, embryo rescue or mutagenesis (see below) to generate diversity and produce hybrid plants that would not exist in nature.
Traits that breeders have tried to incorporate into crop plants in the last 100 years include:
1. Increased quality and yield of the crop
2. Increased tolerance of environmental pressures (salinity, extreme temperature, drought)
3. Resistance to viruses, fungi and bacteria
4. Increased tolerance to insect pests
5. Increased tolerance of herbicides

The Yecoro wheat (right) cultivar is sensitive to salinity, plants resulting from a hybrid cross with cultivar W4910 (left) show greater tolerance to high salinity
Before World War II
Intraspecific hybridization within a plant species was demonstrated by Charles Darwin and Gregor Mendel, and was further developed by geneticists and plant breeders. In the early 20th century, plant breeders realized that Mendel's findings on the non-random nature of inheritance could be applied to seedling populations produced through deliberate pollinations to predict the frequencies of different types.In 1908, George Harrison Shull described heterosis, also known as hybrid vigor. Heterosis describes the tendency of the progeny of a specific cross to outperform both parents. The detection of the usefulness of heterosis for plant breeding has led to the development of inbred lines that reveal a heterotic yield advantage when they are crossed. Maize was the first species where heterosis was widely used to produce hybrids.
By the 1920s, statistical methods were developed to analyze gene action and distinguish heritable variation from variation caused by environment. In 1933, another important breeding technique, cytoplasmic male sterility (CMS), developed in maize, was described by Marcus Morton Rhoades. CMS is a maternally inherited trait that makes the plant produce sterile pollen. This enables the production of hybrids without the need for labour intensive detasseling.
These early breeding techniques resulted in large yield increase in the United States in the early 20th century. Similar yield increases were not produced elsewhere until after World War II, the Green Revolution increased crop production in the developing world in the 1960s.
After World War II
Following World War II a number of techniques were developed that allowed plant breeders to hybridize distantly related species, and artificially induce genetic diversity.When distantly related species are crossed, plant breeders make use of a number of plant tissue culture techniques to produce progeny from otherwise fruitless mating. Interspecific and intergeneric hybrids are produced from a cross of related species or genera that do not normally sexually reproduce with each other. These crosses are referred to as Wide crosses. For example, the cereal triticale is a wheat and rye hybrid. The cells in the plants derived from the first generation created from the cross contained an uneven number of chromosomes and as result was sterile. The cell division inhibitor colchicine was used to double the number of chromosomes in the cell and thus allow the production of a fertile line.
Hybrids may also be produced by a technique called protoplast fusion. In this case protoplasts are fused, usually in an electric field. Viable recombinants can be regenerated in culture.
Chemical mutagens like EMS and DMS, radiation and transposons are used to generate mutants with desirable traits to be bred with other cultivars. Classical plant breeders also generate genetic diversity within a species by exploiting a process called somaclonal variation, which occurs in plants produced from tissue culture, particularly plants derived from callus. Induced polyploidy, and the addition or removal of chromosomes using a technique called chromosome engineering may also be used.
It should be noted that with classical breeding techniques, the breeder does not know exactly what genes have been introduced to the new cultivars. Some scientists therefore argue that plants produced by classical breeding methods should undergo the same safety testing regime as genetically modified plants. There have been instances where plants bred using classical techniques have been unsuitable for human consumption, for example the poison solanine was unintentionally increased to unacceptable levels in certain varieties of potato through plant breeding. New potato varieties are often screened for solanine levels before reaching the marketplace.
Modern plant breeding
Modern plant breeding uses techniques of molecular biology to select, or in the case of genetic modification, to insert, desirable traits into plants.Steps of Plant Breeding
Major activities of plant breeding are following;1. Creation variation
2. Selection
3. Evaluation
4. Release
5. Multiplication
6. Distribution of the new variety
Marker assisted selection
Sometimes many different genes can influence a desirable trait in plant breeding. The use of tools such as molecular markers or DNA fingerprinting can map thousands of genes. This allows plant breeders to screen large populations of plants for those that possess the trait of interest. The screening is based on the presence or absence of a certain gene as determined by laboratory procedures, rather than on the visual identification of the expressed trait in the plant.Reverse Breeding and Doubled Haploids (DH)
A method for efficiently producing homozygous plants from a heterozygous starting plant, which has all desirable traits. This starting plant is induced to produce doubled haploid from haploid cells, and later on creating homozygous/doubled haploid plants from those cells. While in natural offspring recombination occurs and traits can be unlinked from each other, in doubled haploid cells and in the resulting DH plants recombination is no longer an issue. There, a recombination between two corresponding chromosomes does not lead to un-linkage of alleles or traits, since it just leads to recombination with its identical copy. Thus, traits on one chromosome stay linked. Selecting those offspring having the desired set of chromosomes and crossing them will result in a final F1 hybrid plant, having exactly the same set of chromosomes, genes and traits as the starting hybrid plant. The homozygous parental lines can reconstitute the original heterozygous plant by crossing, if desired even in a large quantity. An individual heterozygous plant can be converted into a heterozygous variety (F1 hybrid) without the necessity of vegetative propagation but as the result of the cross of two homozygous/doubled haploid lines derived from the originally selected plant.Genetic modification
Genetic modification of plants is achieved by adding a specific gene or genes to a plant, or by knocking out a gene with RNAi, to produce a desirable phenotype. The plants resulting from adding a gene are often referred to as transgenic plants. If for genetic modification genes of the species or of a crossable plant are used under control of their native promoter, then they are called Cisgenic plants. Genetic modification can produce a plant with the desired trait or traits faster than classical breeding because the majority of the plant's genome is not altered.To genetically modify a plant, a genetic construct must be designed so that the gene to be added or removed will be expressed by the plant. To do this, a promoter to drive transcription and a termination sequence to stop transcription of the new gene, and the gene or genes of interest must be introduced to the plant. A marker for the selection of transformed plants is also included. In the laboratory, antibiotic resistance is a commonly used marker: plants that have been successfully transformed will grow on media containing antibiotics; plants that have not been transformed will die. In some instances markers for selection are removed by backcrossing with the parent plant prior to commercial release.
The majority of commercially released transgenic plants, are currently limited to plants that have introduced resistance to insect pests and herbicides. Insect resistance is achieved through incorporation of a gene from Bacillus thuringiensis (Bt) that encodes a protein that is toxic to some insects. For example, the cotton bollworm, a common cotton pest, feeds on Bt cotton it will ingest the toxin and die. Herbicides usually work by binding to certain plant enzymes and inhibiting their action. The enzymes that the herbicide inhibits are known as the herbicides target site. Herbicide resistance can be engineered into crops by expressing a version of target site protein that is not inhibited by the herbicide. This is the method used to produce glyphosate resistant crop plants.
Genetic modification of plants that can produce pharmaceuticals (and industrial chemicals), sometimes called pharmacrops, is a rather radical new area of plant breeding.
Issues and concerns
Modern plant breeding, whether classical or through genetic engineering, comes with issues of concern, particularly with regard to food crops. The question of whether breeding can have a negative effect on nutritional value is central in this respect. Although relatively little direct research in this area has been done, there are scientific indications that, by favoring certain aspects of a plant's development, other aspects may be retarded. A study published in the Journal of the American College of Nutrition in 2004, entitled Changes in USDA Food Composition Data for 43 Garden Crops, 1950 to 1999, compared nutritional analysis of vegetables done in 1950 and in 1999, and found substantial decreases in six of 13 nutrients measured, including 6% of protein and 38% of riboflavin. Reductions in calcium, phosphorus, iron and ascorbic acid were also found. The study, conducted at the Biochemical Institute, University of Texas at Austin, concluded in summary: "We suggest that any real declines are generally most easily explained by changes in cultivated varieties between 1950 and 1999, in which there may be trade-offs between yield and nutrient content."Plant breeders' rights is also a major and controversial issue. Today, production of new varieties is dominated by commercial plant breeders, who seek to protect their work and collect royalties through national and international agreements based in intellectual property rights. The range of related issues is complex. In the simplest terms, critics of the increasingly restrictive regulations argue that, through a combination of technical and economic pressures, commercial breeders are reducing biodiversity and significantly constraining individuals (such as farmers) from developing and trading seed on a regional level. Efforts to strengthen breeders' rights, for example, by lengthening periods of variety protection, are ongoing.
When new plant breeds or cultivars are bred, they must be maintained and propagated. Some plants are propagated by asexual means while others are propagated by seeds. Seed propagated cultivars require specific control over seed source and production procedures to maintain the integrity of the plant breeds results. Isolation is necessary to prevent cross contamination with related plants or the mixing of seeds after harvesting. Isolation is normally accomplished by planting distance but in certain crops, plants are enclosed in greenhouses or cages (most commonly used when producing F1 hybrids.)
Participatory Plant Breeding
The fruits of major advances in agricultural science, such as those from the Green Revolution, have bypassed millions of farmers in developing countries, most of whom operate small farms under unstable and difficult growing conditions. The adoption of new plant varieties by this group has been low, an issue that has challenged scientists, development workers, governments, donors, and all others with a stake in agricultural progress and the fight against poverty. Their response has been the creation of a novel and promising set of research methods collectively known as participatory plant breeding. Participatory means that farmers are more involved in the breeding process and breeding goals are defined by farmers instead of international seed companies with their large-scale breeding programs. Farmer's groups and NGOs, for example, may wish to affirm local people's rights over genetic resources, produce seeds themselves, build farmers' technical expertise, or develop new products for niche markets, like organically grown food.
Participatory Plant Breeding is carried out, for example, in northern Vietnam, where government scientists work with farmers from the Muong people ethnic minority to improve local rice varieties.
List of notable plant breeders
» Gregor Mendel » Luther Burbank» Roger Doucet
» Carol Deppe
» Norman Borlaug
» Colin Wyatt
» Glenn Drowns
» Alan Kapuler
Notes
- Breeding Field Crops. 1995. Sleper and Poehlman. Page 3- Davis, D.R., Epp, M.D., and Riordan, H.D. 2004. Changes in USDA Food Composition Data for 43 Garden Crops, 1950 to 1999. Journal of the American College of Nutrition 23(6):669-682
- Costa-Font, J. and E. Mossialos. 2007. Food Quality and Preference 18(2007):173-182
- PRGA 2008. Participatory Plant Breeding (PPB)
- Sperling et al. 2001. A Framework For Analizing Participatory Plant Breeding Approaches And Results.
- Ceccarelli 2001. Decentralized-Participatory Plant Breeding: Adapting Crops to Environments and Clients
References
- Borem, A.; Miranda, G. V. Melhoramento de Plantas. 5ª. ed. Viçosa: Editora UFV, 2009. v. 1. 543 p.- Borem, A. (Org.). Domesticação e Melhoramento: espécies amazônicas1. ed. Visconde do Rio Branco: Suprema Grafica e Editora, 2009. (in press)v. 1. 588 p.
- Borem, A. (Org.); Caixeta, E. T. (Org.) . Marcadores Moleculares. 2a.. ed. Visconde do Rio Branco: Suprema Grafica e Editora, 2008. v. 1. 532 p.
- Borem, A.; Condori, M.; Miranda, G. V. Mejoramiento de Plantas (in Spanish). 1. ed. Viçosa: Editora UFV, 2008. v. 1. 438 p.
- McCouch, S. 2004. Diversifying Selection in Plant Breeding. PLoS Biol 2(10): e347.
- Briggs, F.N. and Knowles, P.F. 1967. Introduction to Plant Breeding. Reinhold Publishing Corporation, New York.
- Gepts, P. (2002). A Comparison between Crop Domestication, Classical Plant Breeding, and Genetic Engineering. Crop Science 42:1780–1790
- Schouten, Henk J., Frans A. Krens & Evert Jacobsen Do cisgenic plants warrant less stringent oversight? Nature Biotechnology 24, 753 (2006).
- Schouten, Henk J., Frans A Krens & Evert Jacobsen Cisgenic plants are similar to traditionally bred plants: EMBO reports 7, 750 - 753 (2006).
- Sun, C. et al. 1998. From indica and japonica splitting in common wild rice DNA to the origin and evolution of Asian cultivated rice. Agricultural Archaeology 1998:21-29
Support our developers




