Other Plant Hormones
Since the discovery of auxin, a number of other plant-growth regulators have been uncovered; ethylene, gibberellins, cytokinins, abscisic acid, florigen (and photoperiodism), phytochrome, and wound hormone are examples.Ethylene
While ethylene certainly exhibits hormonal properties in its effects on plants, its simple molecular structure and the fact that it is a gas seem incompatible with our concept of hormones. Thus, it may be better to refer to ethylene as a hormone-like substance. Ethylene’s effect on plants and fruits was observed long before it was known to be produced in plants. For instance, it had long been known that unripened bananas ripened rapidly when stored near a gas leak in a warehouse. Further, shade trees located near leaking street lamps took on altered forms.
Ripe fruits produce ethylene, and ethylene hastens ripening. (The latter fact became known in 1934.) A ripened apple placed in a bowl with green apples will, by its release of ethylene, hasten the ripening of its neighbors, which, in turn, increase their own ethylene production. Ethylene plays various roles in plant behavior. It releases potato tuber buds from dormancy, stimulates leaf abscission, influences the flowering of pineapple plants, induces the formation of adventitious roots in stem cuttings, helps germinating seeds push up through the soil, and helps bring about senescence.
Ethylene is produced not only by ripening fruit but also by flowers, seeds, leaves, and roots. If fruits are bruised or cut, ethylene seems to be produced in a surge. Application of IAA stimulates ethylene production sometimes as much as tenfold. When pea seeds germinate, an increase of ethylene causes the cells of the stem tip to produce a tighter bend, thus aiding the seedling in pushing through to the surface.
Ethylene is used commercially to control the ripening of fruit, as during the shipping of green bananas. Because ethylene hastens ripening, there is often the need to slow the process by careful ventilation to remove the gas. The same effect can be achieved by pumping in carbon dioxide, which counteracts the effects of ethylene.
Ethylene is also theorized to hasten senescence, perhaps by either contributing to a decline of metabolic rate or interfering with RNA synthesis or protein manufacture. The argument regarding senescence in both plants and animals is that organisms, or the cells of organisms, are “born to die.” In plants, this process seems to be advanced by ethylene and delayed by auxins, gibberellins, or a combination of both. It is interesting to note that woody perennials seem to challenge the concept of the aging process being normal. These trees seem to have no mechanism to bring growth to a close. Growth occurs in cambium and presumably can continue without limit. Of course, trees do not live forever. They fall prey to infection or fire; or if the bark is so thick as to protect them from infection and fire (as is the case with the giant Sequoias of California), perhaps they grow until they eventually fall under their own weight.
Gibberellins
Gibberellin is a growth-promoting substance first isolated from the fungus Gibberella fujikuroi. This fungus infects rice seedlings, causing them to grow to great height. The infection is called the “foolish seedling disease.”
Scientists in the Western nations did not learn about gibberellin until after the end of World War II, in approximately 1950. As many as fifty gibberellins have thus far been isolated from fungi and from the healthy tissues of higher plants (buds, leaf primordia, immature seeds, fruit tissue, and roots). Gibberellin production in roots is generous, and gibberellin is translocated to other parts of the plant.
The way whereby gibberellins accelerate growth is not the same as with auxins. Whereas auxins are involved in cell elongation, gibberellins affect the rate of mitosis. Gibberellins, therefore, do not affect curvature of the stem. In some kinds of plants, flowering is induced by the application of a gibberellin. Dormancy of buds and seeds can be broken, and some gibberellins make possible the growth of plants at lower than normal temperatures. A lawn can be induced to grow two to three weeks earlier than is typical by the use of gibberellins. Gibberellins can also be used to increase the size of seedless Grapes. A drop of a gibberellin extract placed on an unfolding leaf will cause the leaf to increase in length by as much as 400 percent. Gibberellins are designated as Al, A2 , and so on. Gibberellic acid, extracted from Gibberella fujikuroi, is designated as A3.
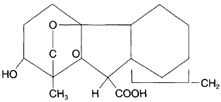 |
| Figure 29-12 Gibberellic acid |
Cytokinins
Growing cells in a tissue culture medium composed in part of coconut milk led to the realization that some substance in coconut milk promotes cell division. The “milk’ of the coconut is actually a liquid endosperm containing large numbers of nuclei. It was from kernels of corn, however, that the substance was first isolated in 1964, twenty years after its presence in coconut milk was known. The substance obtained from corn is called zeatin, and it is one of many cytokinins.
While cytokinin acts mainly to promote cell division, this hormone also serves other functions: cell enlargement in young leaves, tissue differentiation, flowering, fruiting, and delay of aging in leaves. There are now known to be as many as 100 cytokinins, some of which occur naturally and others of which are manufactured. Zeatin is the most active of the natural cytokinins and less active than some of the synthetic ones.
When several living cells are isolated from a living plant and cultured in a medium containing both cytokinin and auxin, cell division proceeds, forming a mass of undifferentiated cells, called a callus. The callus is then able to differentiate and produce both shoot and root, thus developing an entire new plant. Auxin favors root formation, and cytokinin favors the growth of the shoot. This means of vegetative propagation makes possible the perpetuation of superior forms.
Abscisic Acid
Abscisic acid (ABA) is a powerful hormone that inhibits the action of auxins and gibberellins. It is called a stress hormone because it is formed when the plant is subjected to an unfavorable environment. When water is in short supply, abscisic acid contributes to closing the stomates. The presence of the acid in guard cells causes potassium ions to leave the cells; the guard cells thus lose water, and the stomates close. Abscisic acid affects seed dormancy as well as fruit and leaf abscission. Because the action of abscisic acid appears to oppose that of auxins and gibberellins, these hormones are said to be antagonistic.
Photoperiodism and Florigen
Much evidence suggests the existence of a hormone that induces a plant to produce flowers. The discovery of such a flowering hormone has proved elusive, however.
What causes a plant to produce flowers? Some plants produce flowers in the spring, some in the summer, some in the fall. In order for a plant to flower, it must reach a certain maturity. Temperature can also be significant. In many species, the number of hours of daylight or of uninterrupted darkness is the controlling factor. The cocklebur, for example, requires not less than 8½ hours of an uninterrupted darkness in order to produce flowers. If this period of darkness is interrupted by the merest flash of light, flowering will not occur. The wavelength of the interrupting light is significant, however. If the light is of the far-red end of the spectrum, exposure will not inhibit flowering. In fact, exposure to far-red light can reverse the adverse effect of a flash of ordinary light. Thus, if a cocklebur plant is left in darkness for 8½ hours and then brought into ordinary right, flowering will occur. If the dark period is interrupted by a flash of ordinary light, flowering will not occur. If the dark period is interrupted by ordinary light followed by a brief flash of far-red light, however, flowering will occur. The far-red light nullifies the adverse effect of ordinary light and can do so a number of times. If the plant is exposed to ordinary light followed by far-red light followed by ordinary light followed by far-red light, flowering will occur. Whatever light is used last in a series, then, controls whether or not the plant will flower. Light can alter a chemical reaction only if absorbed, and the effect of absorbed far-red light offsets the damage inflicted by absorbed ordinary light.
It is postulated that darkness or far-red light changes an unknown substance from an inhibitory form to a noninhibitory form. It is clear that such a substance is produced in the leaves of the plant because when the leaves are removed after 8½ hours of darkness, flowering does not occur. The substance, then, must be transported from the leaves to the site of flowering. Flowers are modified leaves; thus, the so-called flowering hormone causes the plant to stop producing ordinary leaves and instead produce the modified leaves called flowers.
Although the flowering hormone has not been isolated, it has been given a name: florigen. Several steps are involved in the synthesis and action of the flowering hormone:
- a buildup of florigen achieved by photosynthesis.
- conversion of florigen to a noninhibitory form.
- the synthesis of florigen, also achieved in the dark.
- possibly, a chemical step requiring light
- movement of florigen from the leaves to the site of flowering.
Phytochrome also plays a role in flowering. Its actions are similar to those of florigen, but the interconversions of phytochrome do not alone account for flowering.
Phytochrome is a pale-blue, proteinaceous pigment found in all higher plants. Only minute amounts are produced. Given that phytochrome is essentially invisible, it is not surprising that it remained hidden for so long. Phytochrome was isolated and identified in 1959. A special pigment-analysis instrument had to be utilized to detect it. Phytochrome occurs mostly in meristematic tissue and takes two forms: phytochrome-red (Pr) and phytochrome-far red (Pfr). As mentioned earlier, far red lies at the nearly invisible, long-wave end of the spectrum. Either form of phytochrome can be converted to the other. Phytochrome-far red becomes phytochrome-red when it absorbs far-red light. Conversely, Pr becomes Pfr when it absorbs red light. In nature, Pr becomes Pfr far more than the reverse. Phytochrome-far red converts back to phytochrome-red in the dark. The conversion process in the light is instantaneous. (This should be no surprise given the previous observations regarding the flowering of cocklebur.)
Wound Hormone
Wound hormone is another yet to be isolated hormone. While there is evidence of its existence, the wound hormone remains hidden. Most vascular plants respond to wounding by resuming growth and forming a region of callus (undifferentiated parenchyma cells) covering the wounded area. Wounding triggers the capacity of cells that have ceased dividing to begin growth again. Experiments indicate that the damaged cells produce substances that stimulate cells to resume mitoses. If a wound is immediately washed with water, callus formation does not take place; if wound juice is instead applied, growth will renew.
Support our developers




