Chromosome Translocations
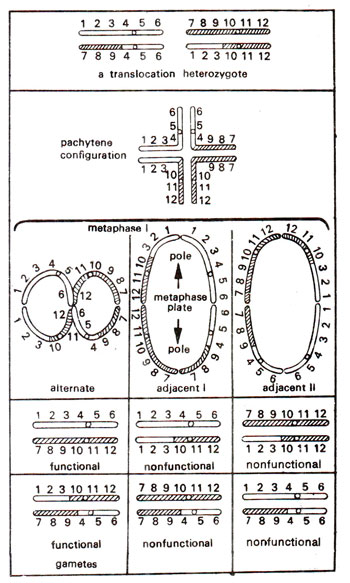
Fig. 19.11. Chromosome pairing and different kinds of gametes formed in a translocation heterozygote.
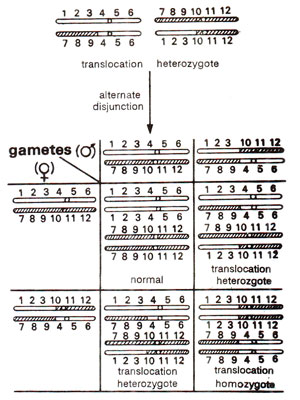
Fig. 19.12. Different kinds of progenies obtained due to self fertilization in a translocation heterozygote.
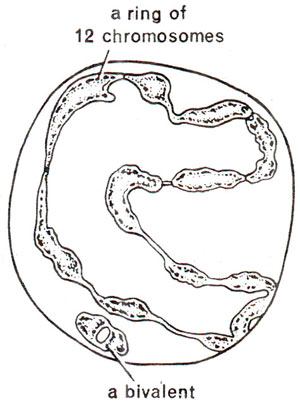
Fig. 19.13. Chromosome complex in Oneothera lamarckiana (2n = 14) showing formation of a ring of 12 chromosomes and a bivalent, as an evidence of multiple interchanges.
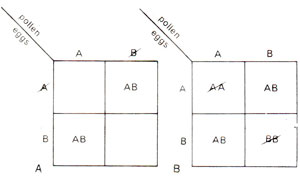
Fig. 19.14. Gametic lethality (A), and zygotic lethality (B) showing balanced lethal systems.
Cytology of a translocation heterozygote
If a translocation is present in one of the two sets of chromosomes, this will be a translocation heterozygote. In such a plant, normal pairing into bivalents will not be possible among chromosomes involved in translocation. Due to pairing between homologous segments of chromosomes, a cross-shaped (+) figure involving four chromosomes will be observed at pachytene (Fig. 19.11). This ring of four chromosomes at metaphase I can have one of the following three orientations :
Alternate. In alternate orientation, alternate chromosomes will be oriented towards the same pole. In other words, adjacent chromosomes will orient towards opposite poles. This will be possible by formation of a figure of eight (Fig. 19.11).
Fig. 19.11. Chromosome pairing and different kinds of gametes formed in a translocation heterozygote.
Adjacent I. In adjacent I orientation, adjacent chromosomes having non- homologous centromeres will orient towards the same pole. In other words, chromosomes having homologous centromeres will orient towards opposite poles. A ring of four chromosomes will be observed.
Adjacent II. In adjacent II orientation, adjacent chromosomes having homologous centromeres will orient towards the same pole. A ring of four chromosomes will be observed.
As shown in Figure 19.11, alternate disjunctions will give functional gametes. Adjacent I and adjacent II disjunctions will form gametes, which would carry duplications or deficiencies and as a result would be non-functional or sterile. Therefore, in a plant having a translocation in heterozygous condition, there will be considerable pollen sterility. A ring of four chromosomes, as described above, is found under conditions when a single interchange is found. If two interchanges are involving three non-homologous chromosomes, a ring of six chromosomes is found, and the size of ring can increase with additional interchanges. More than one ring can also be found if two or more interchanges are independently found, each involving two different non-homologous chromosomes.
Breeding behaviour of a translocation heterozygote
Presence of translocation heterozygosity can be detected by presence of semi- sterility and low seed set. This can then be confirmed at meiosis by quadrivalent formation. As shown above only two types of functional gametes are formed which result from alternate disjunction. The functional gametes will give rise to three kinds of progeny (Fig. 19.12) namely : (i) normal, (ii) translocation heterozygote, and (iii) translocation homozygote. These three types would be obtained in 1 : 2 : 1 ratio.
Fig. 19.12. Different kinds of progenies obtained due to self fertilization in a translocation heterozygote.
Subgenus Euoenothera of genus Oenothera has been studied during 1920-1930 and cytogepetic structure leading to evolution in this group was examined. This group has 2n = 14 and all 7 chromosomes of a haploid complement have median centromeres. Different species in the subgenus Euoenothera, can be classified in three groups : (i) First group is represented by species showing bivalents or small rings at meiosis (e.g. O. hookeri, O. grandiflora, O. argillicola). (ii) Second group is represented by species forming rings of various sizes at meiosis indicating the presence of interchanges. These rings are not permanent but are maintained due to their superiority in adaptive value (e.g. O. irrigua). (iii) The third group is represented by those having permanent translocation heterozygosity involving all chromosomes, so that a ring of 14 chromosomes is regularly formed (e.g. O. biennis, O. strigosa, O. parviflora). In O. lamarckiana, a ring of only 12 instead of a ring of 14 chromosomes is observed (Fig. 19.13). These three groups also differ in phenotypes like flower size, etc. and can be identified. The members of third category behave like pure lines and are actually permanent heterozygotes.
Fig. 19.13. Chromosome complex in Oneothera lamarckiana (2n = 14) showing formation of a ring of 12 chromosomes and a bivalent, as an evidence of multiple interchanges.
Fig. 19.14. Gametic lethality (A), and zygotic lethality (B) showing balanced lethal systems.
Support our developers




