Lambda (λ) bacteriophage exhibits one of the most interesting, but intricate cascade circuits for the two alternative pathways
(lytic cycle or
lysogeny), that it can follow after infection. A brief mention of this phage and its mode of infection was earlier made in
Sexuality and Recombination in Bacteria and Viruses, where it was also explained that the temperate phages like lambda can be classified as episomes, since these are dispensible and exist in free as well as integrated states. The phage can infect bacteria and multiply in the normal manner leading to lysis, but this mode can shift to lysogeny where the phage DNA gets integrated with host DNA and stays tnere 'as prophage till again it becomes lytic.
"How are these two states interconnected, and how does one state shift to another" are the subject of regulation of gene expression. A map of lambda (λ)DNA is given in Figjire 36.2, which has early genes in the middle and late genes at both ends of linear DNA. The linear DNA takes a circular shape during infection, so that the late genes from both ends come together in a cluster to form one transcription unit. When the phage infects a host cell, the early and delayed early genes of the phage are expressed irrespective of whether the phage has to follow lytic or lysogenic mode of development. At this stage, a decision about alternative pathways of development is taken. If the late genes are expressed at this stage, lytic cycle sets in,
but if^a
regulator gene (
cI) synthesizing a
repressor is expressed, the late genes can not be expressed and lysogeny sets in (Fig. 36.3). It is interesting to note that repressor protein coded by
cI regulates its own synthesis also by working as an activator, so that if repressor is inactivated, its further synthesis does not take place and the phage is forced to enter a lytic cycle. Mutants in the gene
cI cannot maintain lysogeny, since then the late' genes can not be stopped from being expressed, due to the absence of active repressor.
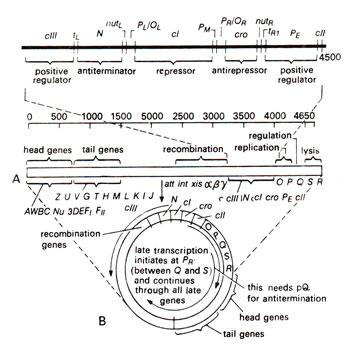
Fig. 36.2. A map of lambda (λ) phage showing clustering of genes (A) and the circularization of DNA during infection (B) thus bringing together the genes on extreme left and extreme right.
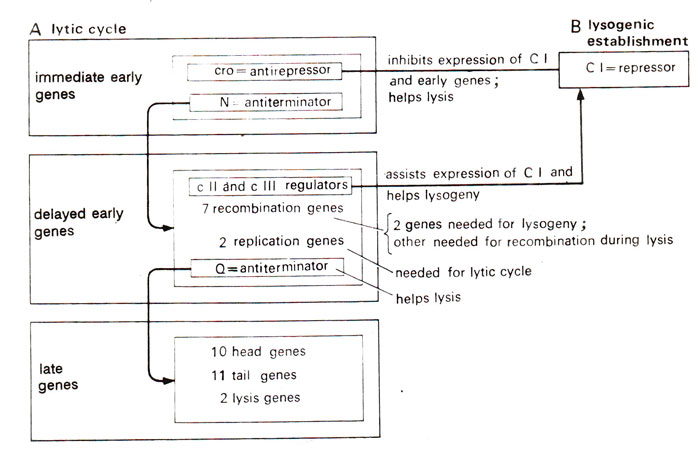
Fig. 36.3. Regulation of the activity of three classes of genes in lytic cycle and in lysogenic establishment in lambda phage (for details see text).
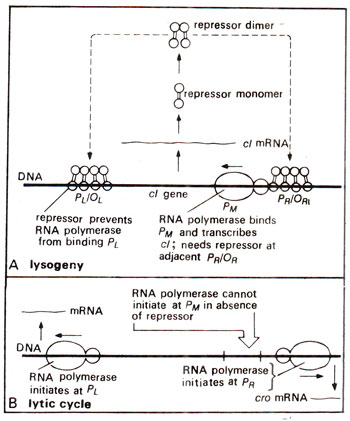
Fig. 36.4. The maintenance of lysogeny by an autogenous circuit of repressor production (A), and the onset of lytic cycle due to interruption in this circuit due to absence of repressor (B).
Three sets of genes in lambda (λ
)phage Immediate early cro and N genes and their control region (PR/OR and PL/OL). As soon as the phage infects, transcription is initiated with the help of two promoters,
PR on the right side and
PL on the left side. The transcription starts on both DNA strands, one strand being transcribed on the left side and the other on the right side. There are two immediate early genes
cro and
N, cro being transcribed on the right side under the control of
PR and
N being transcribed on the left side under the control of
PL. When transcribed,
cro has dual function, namely (i) it prevents synthesis of repressor by
cI (an action which is necessary for lytic cycle), which otherwise causes lysogeny and (ii) it turns off the expression of the immediate early genes including its ownself, since their expression is not needed later in the lytic cycle. Gene
N, as earlier discussed in
Expression of Gene : Protein Synthesis 2. Transcription in Prokaryotes and Eukaryotes,
codes for an antiterminator pN (pN = product of gene N), which allows transcription to proceed into the delayed early genes without the need of any fresh promoter or operator genes. Associated with promoters
PR and
PL are operators
OR and
OL respectively which are sites for repressor binding to prevent RNA polymerase from initiating transcription (Fig. 36.2). The gene
cI (regulator gene coding for repressor) lies in between-
PR/OR and
PL/OL control regions and when active, it not only inhibits the expression of
cro and
N but also promotes its own expression (Fig. 36.4), so that the repressor has a dual function and also works as an
activator for its own
autogenous expression and regulation.
Fig. 36.4. The maintenance of lysogeny by an autogenous circuit of repressor production (A), and the onset of lytic cycle due to interruption in this circuit due to absence of repressor (B).
Delayed early genes (cIIcIII and Q)and their control on lysogeny and lytic cycle. Among the delayed early genes, regulators
cII (on the right) and
cIII (on the left) regulate synthesis of repressor by
cl, to allow phage enter lysogeny (Figs. 36.2, 36.3). On the other hand, a gene
Q (on the right) is a regulator which acts as an
antiterminator, and allows the transcription of late genes meant for lysis. The two pathways (lytic and lysogenic) are so intimately related that it is difficult to predict which pathway will be followed and how is the decision for two alternative pathways is actually taken. This will be further discussed later in this section.
Late genes for lysis (right end) and, tail and head genes (left end). In the phage particle, the late genes are found at the two ends of the linear DNA molecule, genes 5 and
R on the right end, and the tail and head genes on the left end. Genes for recombination are also found on the left side of
cIII and are transcribed as late genes. Soon after the infection, the two ends of DNA molecule join to form a ring, so that all late genes are arranged in a single group containing
S-R genes from right end and head and tail genes
A to
J from left end (Fig. 36.2).
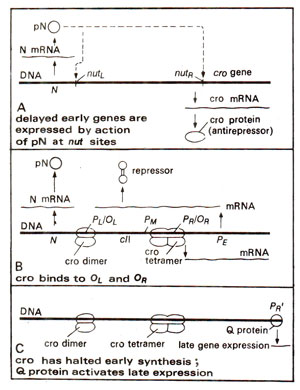
Fig. 36.5. Synthesis and role of Cro protein in prevention of repressor maintenance both through Pm (directly due to binding at Ol and Or) and Pe (indirectly due to switching off the delayed early genes), thus leading to lysis.
There is a promoter gene
PR’ between genes
Q and
S. In the absence of
pQ (product of
Q gene), the transcription from
PR’ is constitutive, but terminates at t
R3 lying close to
PR’ giving a product, 6S RNA (194 bases long). However, when
pQ is present, it suppresses t
R3 and 6S RNA is extended with the result that late genes are heavily expressed. Thus,
Q gene induces transcription of late genes, which continue to be transcribed all through their length. On the left side also, late genes continue to be transcribed. Transcription from both sides stops on the ring molecule, before the RNA polymerase molecules from two sides could clash.
Anti-repressor (product of cro gene) for lytic cycle
We have earlier discussed that
cro gene has dual function of preventing the synthesis of repressor and of turning off the early genes. It codes for a protein (9000 daltons) which forms a dimer and acts on promoter
PM (for
cI gene),
PL (for
N gene on the left side) and
PR (for
cro gene) itself, so that it stops expression of repressor gene
cI as well as that of the early genes (Figs. 36.3, 36.5) including
N on the left and
cro on the right. This fulfils the early requirement for establishment of lytic cycle, because if
cI is expressed, this will stop expression of late genes and thus will establish lysogeny, which is discussed in the next section.
Fig. 36.5. Synthesis and role of Cro protein in prevention of repressor maintenance both through Pm (directly due to binding at Ol and Or) and Pe (indirectly due to switching off the delayed early genes), thus leading to lysis.
Lysogeny through establishment and maintenance of repressor (promoters PE and PM)
In phage lambda (λ)
, a very delicate balance is maintained between lytic cycle and lysogeny, with a very sophisticated and intricate circuit of events, which will be briefly described in this section. It will be seen that the repressor's synthesis is first established with the help of a promoter
PE under the control of genes
cII and
cIII, and is subsequently maintained by the repressor itself through a promoter
PM.
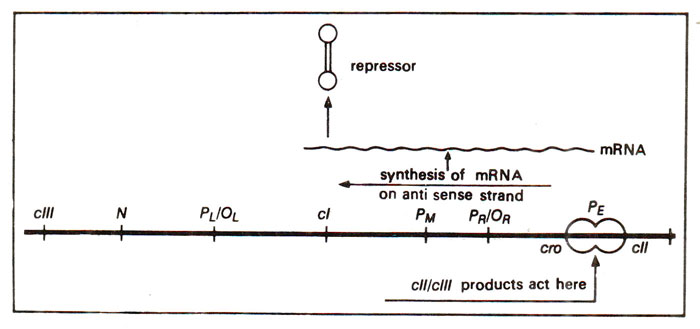
Fig. 36.6. The role of cII and cIII in repressor synthesis due to their action at Pe site leading to transcription of antisense strand.
Promoter PE (or PRE)for repressor establishment and the clear plaque genes (cI, cII, cIII). There is a promoter gene
PE lying between
cro and
cII genes. The product of genes
cII and
cIII act on the promoter
PE, so that the RNA polymerase ' enzyme can initiate transcription towards the left thus synthesizing RNA on genes
cro and
cI. This transcript will consist of (i) the transcript from the antisense strand of
cro gene, which can not be translated (
cro gene is transcribed in the right direction from
PR to yield translatable mRNA) and (ii) mRNA from
cl gene which can be translated to give rise to repressor (Fig. 36.6). The efficiency of synthesis of repressor from
cI through
PE is 7-8 times the synthesis from the promoter
PM (see next section for further details). This repressor synthesized under the control of
PE, binds immediately to the operators
OL and
OR
thus inhibiting transcription from
PL and
PR, leading to turning off the expression of all phage genes. This also halts the synthesis of cII and cIII proteins. Since cII and cIII proteins are unstable and rapidly decay,
PE can not be used any longer for synthesis of repressor. The repressor is now synthesized under the influence of
PM due to the positive control of repressor (already synthesized under the control of promoter
PE), which binds on
OR (which also contains the promoter
PR).
Fig. 36.6. The role of cII and cIII in repressor synthesis due to their action at Pe site leading to transcription of antisense strand.
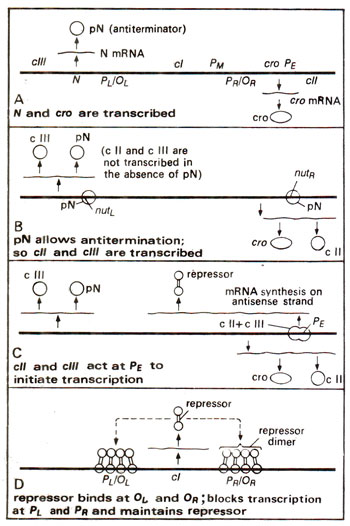
Fig. 36.7. Establishment of lysogeny through a cascade involving the antiterminator protein pN and its further maintenance by the onset of an autogenous repressor maintenance circuit.
The genes
cI, cII and
cIII are so named, because mutants in these genes produce clear plaques, due to the presence of only lysed cells. In the wild type phage, when the lysis occurs, due to the presence of some lysogenic bacteria, clear plaques are not obtained (plaque is a region on the culture medium on the plate, generated due to lysis of bacterial cells). Mutants in
cI (
cI-)differ from the mutants
cII- and
cIII-, since the former can neither establish, nor maintain lysogeny. The mutants
cII- and
cIII- have difficulty in establishing lysogeny, but once established, lysogeny can be maintained. This suggested that the products of genes
cII and
cIII are needed only for the establishment of lysogeny, but not for its maintenance. The genes
cII and
cIII are thus needed to circumvent the difficulty of autogenous circuit of
cI, which can not start the synthesis of repressor
de novo. The different steps involved in establishment and autogenous maintenance of lysogeny are shown in Figure 36.7.
Fig. 36.7. Establishment of lysogeny through a cascade involving the antiterminator protein pN and its further maintenance by the onset of an autogenous repressor maintenance circuit.
Promoter PM (PRM)for autogenous maintenance of repressor. Once the synthesis of repressor is established through the action of
cII and
cIII gene products on
PE , these genes (
cII and
cIII)are switched off by the action of repressor on
OR and
OL. But the repressor works as an activator by binding on
OR, which also contains the promoter
PM (also known
PRM since it is on the right side of
cI)
. By acting on
OR, the repressor helps RNA polymerase to bind on
PM and start further synthesis of repressor. In a remarkable manner it also controls the concentration of repressor, so that when repressor concentration is too high, its further synthesis can be stopped. This is discussed in the following section.
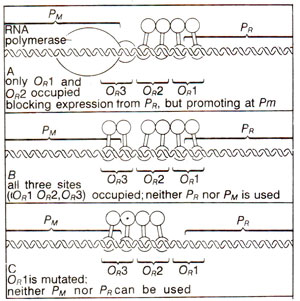
Fig. 36.8. The role of three parts of Or in autogenous regulation of repressor synthesis (for details see text).
Autoregulation of repressor synthesis by cooperative binding of repressor. Each operator (
OR and
OL)has three binding sites,
OR with O
R1-O
R2-O
R3and
OL with O
Ll-O
L2-O
L3
. In each case, site 1 is closest to its promoter (
PR and
PL)and site 3 is farthest. Site 1 has greater affinity (roughly tenfold) for repressor than other sites in each operator, so that the repressor first binds to
OR1and
OL1
. The repressor functions as dimers for maintenance of lysogeny, and at low concentration it binds only to sites
OR1and
OL1
, while with increasing concentration, the repressor dimers can also bind to sites
2(
OR2 and
OL2)and
3(
OR3 and
OL3)
. Presence of repressor on site 1 increases its affinity for site 2 but this interaction does not proceed to site 3. When lysogeny is established, only sites 1 and 2 are occupied but site 3 is not occupied. Since
PR lies within
OR1and
PL lies within
OL1
, occupancy of
OR1
-OR2 and
OL1
-OL2 physically block
PR and
PL respectively and inhibit the expression of all genes thus establishing lysogeny.
The relationship of
OR and
PM is such that
PM (site for RNA polymerase binding) lies rather close to
OR2. When dimers are bound to
OL1
-O,2, the dimer at
OR2 interacts with RNA polymerase in such a way that the latter can utilize
PM for transcription (Fig. 36.8). It is interesting that a repressor dimer bound on
OR2 can inhibit transcription from
PR, but promote transcription from
PM.
Fig. 36.8. The role of three parts of Or in autogenous regulation of repressor synthesis (for details see text).
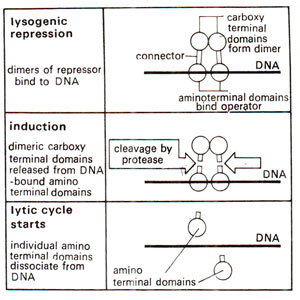
Fig. 36.9. The role of C-terminal domains in controlling the binding of repressor dimers to the operator site (through the N-terminal domain of repressor). (Note that due to cleavage of C-terminal domains, the N-terminal domains lose their affinity for binding).
However, when repressor concentration increases to such an extent, that it may now occupy even
IR3 in addition to
OR1
-OR2, the transcription from
cI is prevented. This effect depends on the fact that
PM lies within
OR3 and when
OR3 is occupied by repressor, it is not available for RNA polymerase. When repressor synthesis stops due to occupancy of
OR3 by repressor, the concentration of repressor consequently falls, and
OR3 again becomes available for RNA polymerase and synthesis of repressor starts once again due to the presence of repressor on
OR2. Thus repressor is an autogenous regulator of its own synthesis causing the inhibition by negative control at its high concentration and promoting by positive control at its low concentration.
Change from lysogeny to lytic cycle by cleavage of repressor dimers in the connector region. The repressor functions as a dimer and each unit (27,000 daltons) has (i) a N-terminal domain (1-92 residues), which binds to operator; (ii) a C-terminal end (133-132 residues), which enhances the affinity of N-terminal domain for binding and (iii) a connector region (93-132 residues). The induction of a lysogenic prophage to enter the lytic cycle is caused by cleavage of repressor in the connector region between 111 and 112 amino acid residues. The cleavage releases the C-terminal domains, so that N-terminal domains now do not have sufficient affinity to remain attached to the operator. N-terminal domains thus dissociate, leading to lytic infection, because now the genes repressed earlier due to repressor, will be expressed causing lysis (Fig. 36.9). This transition from lysogeny to lysis, involves the expression of
Q gene with the help of promoter
PR' giving its product
pQ regulator which helps in the expression of late genes
S and
R (Fig. 36.5).
Fig. 36.9. The role of C-terminal domains in controlling the binding of repressor dimers to the operator site (through the N-terminal domain of repressor). (Note that due to cleavage of C-terminal domains, the N-terminal domains lose their affinity for binding).




