Phylum Nematoda: Roundworms
Phylum Nematoda: Roundworms
Approximately 12,000 species of Nematoda (nem-a-to´da) (Gr., nematos, thread) have been named, but it has been estimated that if all species were known, the number would be nearer 500,000. They live in the sea, in fresh water, and in soil, from polar regions to the tropics, and from mountaintops to the depths of the sea. Good topsoil may contain billions of nematodes per acre. Nematodes also parasitize virtually every type of animal and many plants. Effects of nematode infestation on crops, domestic animals, and humans make this phylum one of the most important of all parasitic animal groups.
Free-living nematodes feed on bacteria, yeasts, fungal hyphae, and algae. They may be saprozoic or coprozoic (live in fecal material). Predatory species may eat rotifers, tardigrades, small annelids, and other nematodes. Many species feed on plant juices from higher plants, which they penetrate, sometimes causing agricultural damage of great proportions. Nematodes themselves may be prey for mites, insect larvae, and even nematode-capturing fungi. Caenorhabditis elegans, a free-living nematode, is easy to culture in the laboratory and has become an invaluable model for studies of basic developmental biology.
Virtually every species of vertebrate and many invertebrates serve as hosts for one or more types of parasitic nematodes. Nematode parasites in hu-mans cause much discomfort, disease, and death, and in domestic animals they are a source of great economic loss.
Form and Function
Distinguishing characteristics of this large group of animals are their cylindrical shape; their flexible, nonliving cuticle; their lack of motile cilia or flagella (except in one species); the muscles of their body wall, which have several unusual features, such as running in a longitudinal direction only, and eutely. Correlated with their lack of cilia, nematodes do not have protonephridia; their excretory system consists of one or more large gland cells opening by an excretory pore, or a canal system without gland cells, or both cells and canals together. Their pharynx is characteristically muscular with a triradiate lumen and resembles the pharynx of gastrotrichs and of kinorhynchs. Use of the pseudocoel as a hydrostatic organ is highly developed in nematodes, and much functional morphology of nematodes can be best understood in the context of the high hydrostatic pressure (turgor) in the pseudocoel.
Most nematode worms are less than 5 cm long, and many are microscopic, but some parasitic nematodes are more than 1 m in length.
The outer body covering is a relatively thick, noncellular cuticle secreted by the underlying epidermis (hypodermis). The hypodermis is syncytial, and its nuclei are located in four hypodermal cords that project inward (Figure 15-10). Dorsal and ventral hypodermal cords bear longitudinal dorsal and ventral nerves, and the lateral cords bear excretory canals. The cuticle is of great functional importance to the worm, serving to contain the high hydrostatic pressure exerted by fluid in the pseudocoel. The several layers of the cuticle are primarily of collagen, a structural protein also abundant in vertebrate connective tissue. Three of the layers are composed of crisscrossing fibers, which confer some longitudinal elasticity on the worm but severely limit its capacity for lateral expansion.
Body wall muscles of nematodes are very unusual. They lie beneath the hypodermis and contract longitudinally only. There are no circular muscles in the body wall. The muscles are arranged in four bands, or quadrants, marked off by the four hypodermal cords (Figure 15-10). Each muscle cell has a contractile fibrillar portion (or spindle) and a noncontractile sarcoplasmic portion (cell body). The spindle is distal and abuts the hypodermis, and the cell body projects into the pseudocoel. The spindle is striated with bands of actin and myosin, reminiscent of vertebrate skeletal muscle (see Figure 9-7). The cell bodies contain the nuclei and are a major depot for glycogen storage in the worm. From each cell body a process or muscle arm extends either to the ventral or the dorsal nerve. Though not unique to nematodes, this arrangement is very curious; in most animals nerve processes (axons) extend to the muscle, rather than the other way around.
The fluid-filled pseudocoel, in which the internal organs lie, constitutes a hydrostatic skeleton. Hydrostatic skeletons, found in many invertebrates, lend support by transmitting the force of muscle contraction to the enclosed, noncompressible fluid. Normally, muscles are arranged antagonistically, so that movement is effected in one direction by contraction of one group of muscles, and movement back in the opposite direction is effected by the antagonistic set of muscles. However, nematodes do not have circular body wall muscles to antagonize the longitudinal muscles; therefore the cuticle must serve that function. As muscles on one side of the body contract, they compress the cuticle on that side, and the force of the contraction is transmitted (by the fluid in the pseudocoel) to the other side of the nematode, stretching the cuticle on that side. This compression and stretching of the cuticle serve to antagonize the muscle and are the forces that return the body to resting position when the muscles relax; this action produces the characteristic thrashing motion seen in nematode movement. An increase in efficiency of this system can be achieved only by an increase in hydrostatic pressure. Consequently, hydrostatic pressure in the nematode pseudocoel is much higher than is usually found in other kinds of animals that have hydrostatic skeletons but that also have antagonistic muscle groups.
The alimentary canal of nematodes consists of a mouth (Figure 15-10), a muscular pharynx, a long nonmuscular intestine, a short rectum, and a terminal anus. Food is sucked into the pharynx when the muscles in its anterior portion contract rapidly and open the lumen. Relaxation of the muscles anterior to the food mass closes the lumen of the pharynx, forcing the food posteriorly toward the intestine. The intestine is one cell-layer thick. Food matter moves posteriorly by body movements and by additional food being passed into the intestine from the pharynx. Defecation is accomplished by muscles that simply pull the anus open, and expulsive force is provided by the high pseudocoelomic pressure that surrounds the gut.
Adults of many parasitic nematodes have an anaerobic energy metabolism; thus, a Krebs cycle and cytochrome system characteristic of aerobic metabolism are absent. They derive energy through glycolysis and probably through some incompletely known electron-transport sequences. Interestingly, some free-living nematodes and free-living stages of parasitic nematodes are obligate aerobes and have a Krebs cycle and cytochrome system.
A ring of nerve tissue and ganglia around the pharynx give rise to small nerves to the anterior end and to two nerve cords, one dorsal and one ventral. Sensory papillae are concentrated around the head and tail. The amphids are a pair of somewhat more complex sensory organs that open on each side of the head at about the same level as the cephalic circle of papillae. The amphidial opening leads into a deep cuticular pit with sensory endings of modified cilia. Amphids are usually reduced in nematode parasites of animals, but most parasitic nematodes bear a bilateral pair of phasmids near the posterior end. They are rather similar in structure to amphids.
Most nematodes are dioecious.
Males are smaller than females, and
their posterior end usually bears a
pair of copulatory spicules (Figure
15-11). Fertilization is internal,
and eggs are usually stored in the
uterus until deposition. After embryonation
a juvenile worm hatches. The
four juvenile stages are each separated
by a molt, or shedding, of the
cuticle. Many parasitic nematodes
have free-living juvenile stages. Others
require an intermediate host to
complete their life cycles.
Some Nematode Parasites
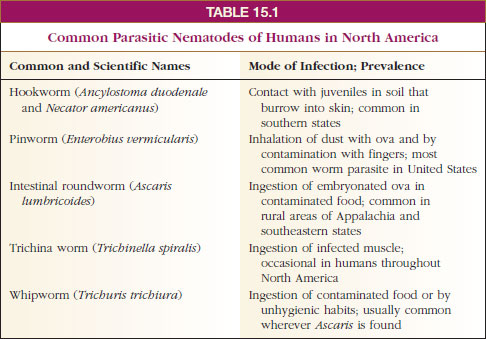
As mentioned previously, nearly all vertebrates and many invertebrates are parasitized by nematodes. A number of these are very important pathogens of humans and domestic animals. A few nematodes are common in humans in North America (Table 15-1), but they and many others usually abound in tropical countries. Space permits mention of only a few in this discussion.
Ascaris lumbricoides: The Large Roundworm of Humans
Because of its size and availability, Ascaris (Gr., askaris, intestinal worm) is usually selected as a type for study in zoology, as well as in experimental work. Thus it is probable that parasitologists know more about structure, physiology, and biochemistry of Ascaris than of any other nematode. This genus includes several species. One of the most common, A. megalocephala, is found in the intestine of horses. Ascaris lumbricoides (Figure 15-12) is one of the most common parasites found in humans; recent surveys have shown a prevalence of up to 64% in some areas of the southeastern United States, and more than 1.2 billion people are infected worldwide. The large roundworm of pigs, A. suum, is morphologically close to A. lumbricoides, and they were long considered the same species.
A female Ascaris may lay 200,000 eggs a day, passing out in the host’s feces. Given suitable soil conditions, embryonation is complete within 2 weeks. Direct sunlight and high temperatures are rapidly lethal, but the eggs have an amazing tolerance to other adverse conditions, such as desiccation or lack of oxygen. Shelled juveniles can remain viable for many months or even years in the soil. Infection usually occurs when eggs are ingested with uncooked vegetables or when children put soiled fingers or toys in their mouths. Unsanitary defecation habits “seed” the soil, and viable eggs remain long after all signs of the fecal matter have disappeared.
When a host swallows embryonated eggs, the tiny juveniles hatch. They burrow through the intestinal wall into veins or lymph vessels and are carried through the heart to the lungs. There they break out into the alveoli and are carried up to the tracheae. If the infection is large, they may cause a serious pneumonia at this stage. On reaching the pharynx, juveniles are swallowed, passed through the stomach, and finally mature about 2 months after the eggs were ingested. In the intestine, where they feed on intestinal contents, the worms cause abdominal symptoms and allergic reactions, and in large numbers they may cause intestinal blockage. Perforation of the intestine with resultant peritonitis is not uncommon, and wandering worms may occasionally emerge from the anus or throat or may enter the trachea or eustachian tubes and middle ears.
Hookworms
Hookworms are so named because the anterior end curves dorsally, suggesting a hook. The most common species is Necator americanus (L. necator, killer), whose females are up to 11 mm long. Males can reach 9 mm in length. Large plates in their mouths (Figure 15-13) cut into the intestinal mucosa of the host where they suck blood and pump it through their intestine, partially digesting it and absorbing the nutrients. They suck much more blood than they need for food, and heavy infections cause anemia in patients. Hookworm disease in children may result in retarded mental and physical growth and a general loss of energy.
Eggs pass in the feces, and juveniles hatch in the soil, where they live on bacteria. When human skin comes in contact with infected soil, infective juveniles burrow through the skin to the blood, and reach the lungs and finally the intestine in a manner similar to that described for Ascaris.
Trichina Worm
Trichinella spiralis (Gr. trichinos, of
hair, + -ella, diminutive) is one of
the species of tiny nematodes
responsible for the potentially lethal
disease trichinosis. Adult worms burrow
in the mucosa of the small intestine
where females produce living
young. Juveniles penetrate blood vessels
and are carried throughout the
body, where they may be found in
almost any tissue or body space.
Eventually, they penetrate skeletal
muscle cells, becoming one of the
largest known intracellular parasites.
Juveniles cause astonishing redirection
of gene expression in their host
cell, which loses its striations and
becomes a nurse cell that nourishes
the worm (Figure 15-14). When meat
containing live juveniles is swallowed,
the worms are liberated into
the intestine where they mature.
Trichinella spp. can infect a wide variety of mammals in addition to humans, including hogs, rats, cats, and dogs. Hogs become infected by eating garbage containing pork scraps with juveniles or by eating infected rats. In addition to T. spiralis, we now know there are four other sibling species in the genus. They differ in geographic distribution, infectivity to different host species, and freezing resistance.
Heavy infections may cause death, but lighter infections are much more common—about 2.4% of the population of the United States is infected.
Pinworms
Pinworms, Enterobius vermicularis (Gr. enteron, intestine, + bios, life), cause relatively little disease, but they are the most common helminth parasites in the United States, estimated at 30% in children and 16% in adults. Adult parasites (Figure 15-15) live in the large intestine and cecum. Females, up to about 12 mm in length, migrate to the anal region at night to lay their eggs (Figure 15-15). Scratching the resultant itch effectively contaminates hands and bedclothes. Eggs develop rapidly and become infective within 6 hours at body temperature. When they are swallowed, they hatch in the duodenum, and the worms mature in the large intestine.
Members of this order of nematodes have haplodiploidy, a characteristic shared with a few other animal groups, notably many hymenopteran insects. Males are haploid and are produced parthenogenetically; females are diploid and arise from fertilized eggs.
Filarial Worms
At least eight species of filarial nematodes infect humans, and some of these are major causes of disease. Some 250 million people in tropical countries are infected with Wuchereria bancrofti (named for Otto Wucherer) or Brugia malayi (named for S. L. Brug), which places these species among the scourges of humanity. The worms live in the lymphatic system, and females are as long as 100 mm. The disease symptoms are associated with inflammation and obstruction of the lymphatic system. Females release live young, tiny microfilariae, into the blood and lymphatic system. As they feed, mosquitos ingest microfilariae, and they develop in mosquitos to the infective stage. They escape from the mosquito when it is feeding again on a human and penetrate the wound made by the mosquito bite.
The dramatic manifestations of elephantiasis are produced occasionally after long and repeated exposure to the worms. The condition is marked by an excessive growth of connective tissue and enormous swelling of affected parts, such as the scrotum, legs, arms, and more rarely, the vulva and breasts (Figure 15-16).
Another filarial worm causes river blindness (onchocerciasis) and is carried by black flies. It infects more than 30 million people in parts of Africa, Arabia, Central America, and South America.
The most common filarial worm in
the United States is probably the dog
heartworm, Dirofilaria immitis (Figure
15-17). Carried by mosquitos, it
also can infect other canids, cats, ferrets,
sea lions, and occasionally humans. Along the Atlantic and Gulf
Coast states and northward along the
Mississippi River throughout the midwestern
states, prevalence in dogs is
up to 45%. It occurs in other states at a
lower prevalence. This worm causes a
very serious disease among dogs, and
no responsible owner should fail to
provide “heartworm pills” for a dog
during mosquito season.
Classification of Phylum Nematoda
Classification of nematodes is somewhat
more satisfactory at the order
and superfamily level; division into
classes relies on characteristics that
are not striking and that are difficult
for novices to distinguish. The classification
given here is that proposed
by Adamson,* whose analysis indicated
that the traditional class Aphasmidia
was paraphyletic.
Class Rhabditea (rab-di´te-a) (Gr. rhabdos, a rod) Amphids ventrally coiled or derived therefrom; three esophageal glands; some with phasmids; both free-living and parasitic forms. Examples: Caenorhabditis, Ascaris, Enterobius, Necator, Wuchereria.
Class Enoplea (ee-no´ple-a) (Gr. enoplos, armed) Amphids generally well-developed, pocket-like; five or more esophageal glands; phasmids absent; excretory system lacking lateral canals, formed of single, ventral, glandular cells, or entirely absent; mostly free living, but includes some parasites. Examples: Dioctophyme, Trichinella, Trichuris.
* Adamson, M. 1987. Canad. J. Zool. 65:1478–1482.
Approximately 12,000 species of Nematoda (nem-a-to´da) (Gr., nematos, thread) have been named, but it has been estimated that if all species were known, the number would be nearer 500,000. They live in the sea, in fresh water, and in soil, from polar regions to the tropics, and from mountaintops to the depths of the sea. Good topsoil may contain billions of nematodes per acre. Nematodes also parasitize virtually every type of animal and many plants. Effects of nematode infestation on crops, domestic animals, and humans make this phylum one of the most important of all parasitic animal groups.
Free-living nematodes feed on bacteria, yeasts, fungal hyphae, and algae. They may be saprozoic or coprozoic (live in fecal material). Predatory species may eat rotifers, tardigrades, small annelids, and other nematodes. Many species feed on plant juices from higher plants, which they penetrate, sometimes causing agricultural damage of great proportions. Nematodes themselves may be prey for mites, insect larvae, and even nematode-capturing fungi. Caenorhabditis elegans, a free-living nematode, is easy to culture in the laboratory and has become an invaluable model for studies of basic developmental biology.
Virtually every species of vertebrate and many invertebrates serve as hosts for one or more types of parasitic nematodes. Nematode parasites in hu-mans cause much discomfort, disease, and death, and in domestic animals they are a source of great economic loss.
Form and Function
Distinguishing characteristics of this large group of animals are their cylindrical shape; their flexible, nonliving cuticle; their lack of motile cilia or flagella (except in one species); the muscles of their body wall, which have several unusual features, such as running in a longitudinal direction only, and eutely. Correlated with their lack of cilia, nematodes do not have protonephridia; their excretory system consists of one or more large gland cells opening by an excretory pore, or a canal system without gland cells, or both cells and canals together. Their pharynx is characteristically muscular with a triradiate lumen and resembles the pharynx of gastrotrichs and of kinorhynchs. Use of the pseudocoel as a hydrostatic organ is highly developed in nematodes, and much functional morphology of nematodes can be best understood in the context of the high hydrostatic pressure (turgor) in the pseudocoel.
Most nematode worms are less than 5 cm long, and many are microscopic, but some parasitic nematodes are more than 1 m in length.
The outer body covering is a relatively thick, noncellular cuticle secreted by the underlying epidermis (hypodermis). The hypodermis is syncytial, and its nuclei are located in four hypodermal cords that project inward (Figure 15-10). Dorsal and ventral hypodermal cords bear longitudinal dorsal and ventral nerves, and the lateral cords bear excretory canals. The cuticle is of great functional importance to the worm, serving to contain the high hydrostatic pressure exerted by fluid in the pseudocoel. The several layers of the cuticle are primarily of collagen, a structural protein also abundant in vertebrate connective tissue. Three of the layers are composed of crisscrossing fibers, which confer some longitudinal elasticity on the worm but severely limit its capacity for lateral expansion.
Body wall muscles of nematodes are very unusual. They lie beneath the hypodermis and contract longitudinally only. There are no circular muscles in the body wall. The muscles are arranged in four bands, or quadrants, marked off by the four hypodermal cords (Figure 15-10). Each muscle cell has a contractile fibrillar portion (or spindle) and a noncontractile sarcoplasmic portion (cell body). The spindle is distal and abuts the hypodermis, and the cell body projects into the pseudocoel. The spindle is striated with bands of actin and myosin, reminiscent of vertebrate skeletal muscle (see Figure 9-7). The cell bodies contain the nuclei and are a major depot for glycogen storage in the worm. From each cell body a process or muscle arm extends either to the ventral or the dorsal nerve. Though not unique to nematodes, this arrangement is very curious; in most animals nerve processes (axons) extend to the muscle, rather than the other way around.
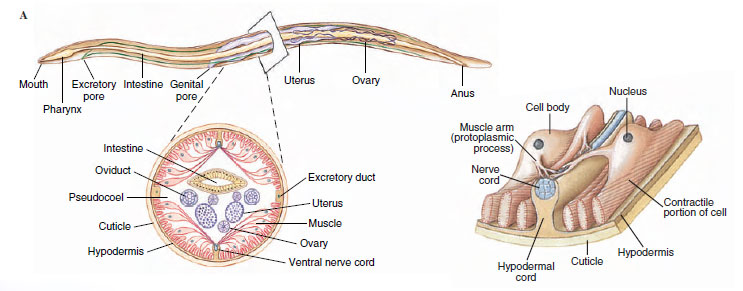 |
| Figure 15-10 A, Structure of a nematode as illustrated by Ascaris female. Ascaris has two ovaries and uteri, which open to the outside by a common genital pore. B, Cross section. C, Single muscle cell; spindle abuts hypodermis, muscle arm extends to dorsal or ventral nerve. |
The fluid-filled pseudocoel, in which the internal organs lie, constitutes a hydrostatic skeleton. Hydrostatic skeletons, found in many invertebrates, lend support by transmitting the force of muscle contraction to the enclosed, noncompressible fluid. Normally, muscles are arranged antagonistically, so that movement is effected in one direction by contraction of one group of muscles, and movement back in the opposite direction is effected by the antagonistic set of muscles. However, nematodes do not have circular body wall muscles to antagonize the longitudinal muscles; therefore the cuticle must serve that function. As muscles on one side of the body contract, they compress the cuticle on that side, and the force of the contraction is transmitted (by the fluid in the pseudocoel) to the other side of the nematode, stretching the cuticle on that side. This compression and stretching of the cuticle serve to antagonize the muscle and are the forces that return the body to resting position when the muscles relax; this action produces the characteristic thrashing motion seen in nematode movement. An increase in efficiency of this system can be achieved only by an increase in hydrostatic pressure. Consequently, hydrostatic pressure in the nematode pseudocoel is much higher than is usually found in other kinds of animals that have hydrostatic skeletons but that also have antagonistic muscle groups.
The alimentary canal of nematodes consists of a mouth (Figure 15-10), a muscular pharynx, a long nonmuscular intestine, a short rectum, and a terminal anus. Food is sucked into the pharynx when the muscles in its anterior portion contract rapidly and open the lumen. Relaxation of the muscles anterior to the food mass closes the lumen of the pharynx, forcing the food posteriorly toward the intestine. The intestine is one cell-layer thick. Food matter moves posteriorly by body movements and by additional food being passed into the intestine from the pharynx. Defecation is accomplished by muscles that simply pull the anus open, and expulsive force is provided by the high pseudocoelomic pressure that surrounds the gut.
Adults of many parasitic nematodes have an anaerobic energy metabolism; thus, a Krebs cycle and cytochrome system characteristic of aerobic metabolism are absent. They derive energy through glycolysis and probably through some incompletely known electron-transport sequences. Interestingly, some free-living nematodes and free-living stages of parasitic nematodes are obligate aerobes and have a Krebs cycle and cytochrome system.
A ring of nerve tissue and ganglia around the pharynx give rise to small nerves to the anterior end and to two nerve cords, one dorsal and one ventral. Sensory papillae are concentrated around the head and tail. The amphids are a pair of somewhat more complex sensory organs that open on each side of the head at about the same level as the cephalic circle of papillae. The amphidial opening leads into a deep cuticular pit with sensory endings of modified cilia. Amphids are usually reduced in nematode parasites of animals, but most parasitic nematodes bear a bilateral pair of phasmids near the posterior end. They are rather similar in structure to amphids.
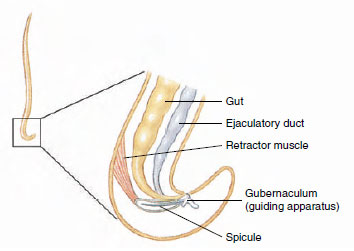 |
| Figure 15-11 Posterior end of a male nematode. |
Some Nematode Parasites
As mentioned previously, nearly all vertebrates and many invertebrates are parasitized by nematodes. A number of these are very important pathogens of humans and domestic animals. A few nematodes are common in humans in North America (Table 15-1), but they and many others usually abound in tropical countries. Space permits mention of only a few in this discussion.
Ascaris lumbricoides: The Large Roundworm of Humans
Because of its size and availability, Ascaris (Gr., askaris, intestinal worm) is usually selected as a type for study in zoology, as well as in experimental work. Thus it is probable that parasitologists know more about structure, physiology, and biochemistry of Ascaris than of any other nematode. This genus includes several species. One of the most common, A. megalocephala, is found in the intestine of horses. Ascaris lumbricoides (Figure 15-12) is one of the most common parasites found in humans; recent surveys have shown a prevalence of up to 64% in some areas of the southeastern United States, and more than 1.2 billion people are infected worldwide. The large roundworm of pigs, A. suum, is morphologically close to A. lumbricoides, and they were long considered the same species.
 |
A female Ascaris may lay 200,000 eggs a day, passing out in the host’s feces. Given suitable soil conditions, embryonation is complete within 2 weeks. Direct sunlight and high temperatures are rapidly lethal, but the eggs have an amazing tolerance to other adverse conditions, such as desiccation or lack of oxygen. Shelled juveniles can remain viable for many months or even years in the soil. Infection usually occurs when eggs are ingested with uncooked vegetables or when children put soiled fingers or toys in their mouths. Unsanitary defecation habits “seed” the soil, and viable eggs remain long after all signs of the fecal matter have disappeared.
When a host swallows embryonated eggs, the tiny juveniles hatch. They burrow through the intestinal wall into veins or lymph vessels and are carried through the heart to the lungs. There they break out into the alveoli and are carried up to the tracheae. If the infection is large, they may cause a serious pneumonia at this stage. On reaching the pharynx, juveniles are swallowed, passed through the stomach, and finally mature about 2 months after the eggs were ingested. In the intestine, where they feed on intestinal contents, the worms cause abdominal symptoms and allergic reactions, and in large numbers they may cause intestinal blockage. Perforation of the intestine with resultant peritonitis is not uncommon, and wandering worms may occasionally emerge from the anus or throat or may enter the trachea or eustachian tubes and middle ears.
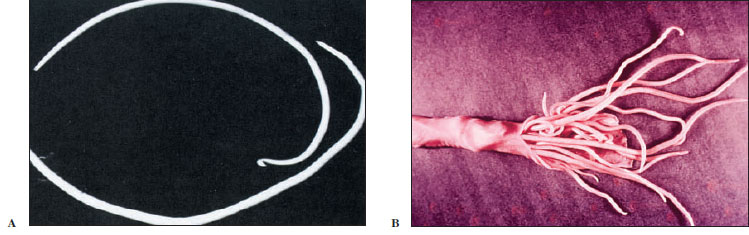 |
| Figure 15-12 A, Intestinal roundworm Ascaris lumbricoides, male and female. Male, top, is smaller and has characteristic sharp kink in the end of the tail. Females of this large nematode may be over 30 cm long. B, Intestine of a pig, nearly completely blocked by Ascaris suum. Such heavy infections are also fairly common with A. lumbricoides in humans. |
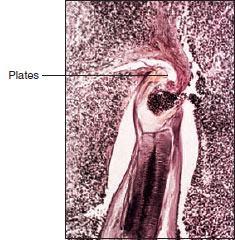 |
| Figure 15-13 Section through anterior end of hookworm attached to dog intestine. Note cutting plates of mouth pinching off mucosa from which the thick muscular pharynx sucks blood. Esophageal glands secrete nticoagulant to prevent blood clotting. |
Hookworms are so named because the anterior end curves dorsally, suggesting a hook. The most common species is Necator americanus (L. necator, killer), whose females are up to 11 mm long. Males can reach 9 mm in length. Large plates in their mouths (Figure 15-13) cut into the intestinal mucosa of the host where they suck blood and pump it through their intestine, partially digesting it and absorbing the nutrients. They suck much more blood than they need for food, and heavy infections cause anemia in patients. Hookworm disease in children may result in retarded mental and physical growth and a general loss of energy.
Eggs pass in the feces, and juveniles hatch in the soil, where they live on bacteria. When human skin comes in contact with infected soil, infective juveniles burrow through the skin to the blood, and reach the lungs and finally the intestine in a manner similar to that described for Ascaris.
Trichina Worm
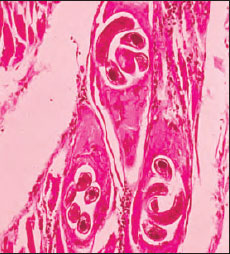 |
| Figure 15-14 Section of muscle infected with trichina worm Trichinella spiralis, human case. The juveniles lie within muscle cells that the worms have induced to transform into nurse cells (commonly called cysts). An inflammatory reaction is evident around the nurse cells. Juveniles may live 10 to 20 years, and nurse cells eventually may calcify. |
Trichinella spp. can infect a wide variety of mammals in addition to humans, including hogs, rats, cats, and dogs. Hogs become infected by eating garbage containing pork scraps with juveniles or by eating infected rats. In addition to T. spiralis, we now know there are four other sibling species in the genus. They differ in geographic distribution, infectivity to different host species, and freezing resistance.
Heavy infections may cause death, but lighter infections are much more common—about 2.4% of the population of the United States is infected.
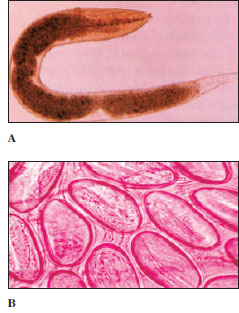 |
| Figure 15-15 Pinworms, Enterobius vermicularis. A, Female worm from human large intestine (slightly flattened in preparation), magnified about 20 times. B, Group of pinworm eggs, which are usually discharged at night around the anus of the host, who, by scratching during sleep, gets fingernails and clothing contaminated. This may be the most common and widespread of all human helminth parasites. |
Pinworms, Enterobius vermicularis (Gr. enteron, intestine, + bios, life), cause relatively little disease, but they are the most common helminth parasites in the United States, estimated at 30% in children and 16% in adults. Adult parasites (Figure 15-15) live in the large intestine and cecum. Females, up to about 12 mm in length, migrate to the anal region at night to lay their eggs (Figure 15-15). Scratching the resultant itch effectively contaminates hands and bedclothes. Eggs develop rapidly and become infective within 6 hours at body temperature. When they are swallowed, they hatch in the duodenum, and the worms mature in the large intestine.
Members of this order of nematodes have haplodiploidy, a characteristic shared with a few other animal groups, notably many hymenopteran insects. Males are haploid and are produced parthenogenetically; females are diploid and arise from fertilized eggs.
Filarial Worms
At least eight species of filarial nematodes infect humans, and some of these are major causes of disease. Some 250 million people in tropical countries are infected with Wuchereria bancrofti (named for Otto Wucherer) or Brugia malayi (named for S. L. Brug), which places these species among the scourges of humanity. The worms live in the lymphatic system, and females are as long as 100 mm. The disease symptoms are associated with inflammation and obstruction of the lymphatic system. Females release live young, tiny microfilariae, into the blood and lymphatic system. As they feed, mosquitos ingest microfilariae, and they develop in mosquitos to the infective stage. They escape from the mosquito when it is feeding again on a human and penetrate the wound made by the mosquito bite.
The dramatic manifestations of elephantiasis are produced occasionally after long and repeated exposure to the worms. The condition is marked by an excessive growth of connective tissue and enormous swelling of affected parts, such as the scrotum, legs, arms, and more rarely, the vulva and breasts (Figure 15-16).
Another filarial worm causes river blindness (onchocerciasis) and is carried by black flies. It infects more than 30 million people in parts of Africa, Arabia, Central America, and South America.
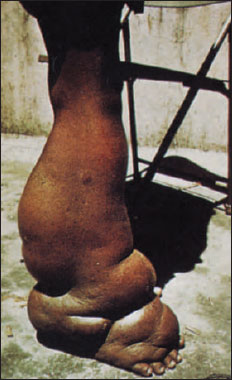 |
| Figure 15-16 Elephantiasis of leg caused by adult filarial worms of Wuchereria bancrofti, which live in lymph passages and block the flow of lymph, Tiny juveniles, called microfilariae, are ingested with blood meal of mosquitos, where they develop to infective stage and are transmitted to a new host. |
Classification of Phylum Nematoda
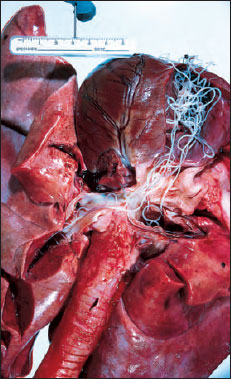 |
| Figure 15-17 Dirofilaria immitis in right ventricle, extending up into right and left pulmonary arteries of an eightyear- old Irish setter. |
Class Rhabditea (rab-di´te-a) (Gr. rhabdos, a rod) Amphids ventrally coiled or derived therefrom; three esophageal glands; some with phasmids; both free-living and parasitic forms. Examples: Caenorhabditis, Ascaris, Enterobius, Necator, Wuchereria.
Class Enoplea (ee-no´ple-a) (Gr. enoplos, armed) Amphids generally well-developed, pocket-like; five or more esophageal glands; phasmids absent; excretory system lacking lateral canals, formed of single, ventral, glandular cells, or entirely absent; mostly free living, but includes some parasites. Examples: Dioctophyme, Trichinella, Trichuris.
* Adamson, M. 1987. Canad. J. Zool. 65:1478–1482.
Support our developers




