Mendel’s First Law
Mendelian Laws
of Inheritance
Mendel’s First Law
Mendel’s law of segregation states that in the formation of gametes, paired factors specifying alternative phenotypes (visible traits) segregate independently of one another. In one of Mendel’s original experiments, he pollinated pure-line tall plants with the pollen of pure-line dwarf plants. Thus the visible characteristics, or phenotypes, of the parents were tall and dwarf. Mendel found that all progeny in the first generation (F1) were tall, just as tall as the tall parents of the cross. The reciprocal cross—dwarf plants pollinated with tall plants—gave the same result. The tall phenotype was observed in progeny no matter which way the cross was made. Obviously, this kind of inheritance was not a blending of two traits, since none of the progeny was of intermediate size.
Next Mendel self-fertilized (“selfed”) the tall F1 plants and raised several hundred progeny, the second (F2) generation. This time, both tall and dwarf plants appeared. Again, there was no blending (no plants of intermediate size), but the appearance of dwarf plants from all tall parental plants was surprising. The dwarf trait, present in the grandparents but not in the parents, had reappeared. When he counted the actual number of tall and dwarf plants in the F2 generation, he discovered that there were almost exactly three times more tall plants than dwarf ones.
Mendel then repeated this experiment for the six other contrasting traits that he had chosen, and in every case he obtained ratios very close to 3:1 (see Figure 5-1). At this point it must have been clear to Mendel that he was dealing with hereditary determinants for the contrasting traits that did not blend when brought together. Even though the dwarf trait disappeared in the F1 generation, it reappeared fully expressed in the F2 generation. He realized that the F1 generation plants carried determinants (which he called “factors”) of both tall and dwarf parents, even though only the tall trait was expressed in the F1 generation.
Mendel called the tall factor dominant and the short recessive. Similarly, the other pairs of traits that he studied showed dominance and recessiveness. Whenever a dominant factor is present, the recessive one cannot produce its effect. The recessive trait will appear only when both factors are recessive, or in other words, in a pure condition.
In representing his crosses, Mendel used letters as symbols; dominant traits were represented by capital letters, and for recessive traits he used the corresponding lowercase letters. Modern geneticists still follow this custom. Thus the factors for pure tall plants might be represented by t/t, the pure recessive by t/t, and the mix, or hybrid, of the two plants by t/t. The slash mark is to indicate that the alleles are on homologous chromosomes. The zygote bears the complete genetic constitution of the organism. All the gametes produced by t/t must necessarily be T, whereas those produced by t/t must be t. Therefore a zygote produced by union of the two must be t/t, or a heterozygote. On the other hand, the pure tall plants (t/t) and pure dwarf plants (t/t) are homozygotes, meaning that the paired factors (alleles) are alike on the homologous chromosomes. A cross involving only one pair of contrasting traits is called a monohybrid cross.
In the cross between tall and dwarf plants there were two phenotypes: tall and dwarf. On the basis of genetic formulas there are three hereditary types: T/T, T/t, and t/t. These are called genotypes. A genotype is an allelic combination (T/T, T/t, or t/t), and the phenotype is the corresponding appearance of the organism (tall or dwarf).
One of Mendel’s original crosses (tall plant and dwarf plant) could be represented as follows:
In other words, all possible combinations
of F1 gametes in the F2 zygotes will
yield a 3:1 phenotypic ratio and a 1:2:1
genotypic ratio. It is convenient in such
crosses to use the checkerboard method
devised by Punnett (Punnett square) for
representing the various combinations
resulting from a cross. In the F2 cross the
following scheme would apply:
The next step was an important one because it enabled Mendel to test his hypothesis that every plant contained nonblending factors from both parents. He self-fertilized the plants in the F2 generation; that is, the stigma of a flower was fertilized by the pollen of the same flower.
The results showed that self-pollinated
F2 dwarf plants produced only
dwarf plants, whereas one-third of the
F2 tall plants produced tall and the other
two-thirds produced both tall and dwarf
in the ratio of 3:1, just as the F1 plants
had done. Genotypes and phenotypes
were as follows:
This experiment showed that the dwarf plants were pure because they at all times gave rise to short plants when self-pollinated; the tall plants contained both pure tall and hybrid tall. It also demonstrated that, although the dwarf trait disappeared in the F1 plants, which were all tall, dwarfness appeared in the F2 plants.
Mendel reasoned that the factors for tallness and dwarfness were units that did not blend when they were together. The F1 generation contained both of these units or factors, but when these plants formed their germ cells, the factors separated so that each germ cell had only one factor. In a pure plant both factors were alike; in a hybrid they were different. He concluded that individual germ cells were always pure with respect to a pair of contrasting factors, even though the germ cells were formed from hybrid individuals possessing both contrasting factors.
This idea formed the basis for his law of segregation, that is, that whenever two factors are brought together in a hybrid, they segregate into separate gametes that are produced by the hybrid. Either one of the paired factors or alleles of the parent pass with equal frequency to the gametes. We now understand that the factors segregate because there are two alleles for the character, one on each chromosome of a homologous pair, but the gametes receive only one of each in meiosis. Thus in current usage the law of segregation refers to the parting of homologous chromosomes during meiosis.
Mendel’s great contribution was his quantitative approach to inheritance. This really marks the birth of genetics, because before Mendel, people believed that traits were blended like mixing together two colors of paint, a notion that unfortunately still lingers in the minds of many and was a problem for Darwin’s theory of natural selection when he first proposed it. If traits were blended, variability would be lost in hybridization between individuals. With particulate inheritance, on the other hand, different variations are retained and can be shuffled about and resorted like blocks.
In not reporting conflicting findings, which must surely have arisen as they do in any original research, Mendel has been accused of “cooking” his results.The chances are, however, that he carefully avoided ambiguous material to strengthen his central message, which we still regard as an exemplary achievement in experimental analysis.
Testcross
When one of the alleles is dominant, heterozygous individuals are identical in phenotype to individuals homozygous for the dominant allele. Therefore you cannot determine the genotypes of these individuals just by observing their phenotypes. For instance, in Mendel’s experiment of tall and dwarf traits, it is impossible to determine the genetic constitution of the tall plants of the F2 generation by mere inspection of the tall plants. Three-fourths of this generation are tall, but which of them are heterozygotes?
As Mendel reasoned, the test is to
cross the questionable individuals with
pure recessives. If the tall plant is
homozygous, all the offspring in such
a testcross are tall, thus:
All of the offspring are T/t (hybrid tall).
If, on the other hand, the tall plant is
heterozygous, half of the offspring are
tall and half dwarf, thus:
The testcross is often used in modern genetics for the analysis of the genetic constitution of the offspring, as well as for a quick way to make desirable homozygous stocks of animals and plants.
Intermediate Inheritance
In some cases neither allele is completely dominant over the other, and the heterozygous phenotype appears either intermediate between or even quite distinct from those of the parents. This is called intermediate inheritance, or incomplete dominance. In the four-o’clock flower (Mirabilis), two allelic variants determine red versus pink or white flowers; homozygotes are red or white flowered,
but heterozygotes
have pink flowers. In a certain
strain of chickens, a cross between
those with black and splashed white
feathers produces offspring that are
not gray but a distinctive color called
Andalusian blue (Figure 5-5). In each
case, if the F1s are crossed, the
F2s have a ratio of 1:2:1 in colors, or
1 red: 2 pink: 1 white in four-o’clock
flowers and 1 black: 2 blue: 1 white for
Andalusian chickens. This can be illustrated
for the chickens as follows:
When neither of the alleles is recessive, it is customary to represent both by capital letters and to distinguish them by the addition of a “prime”sign (B') or by superscript letters, for example,Bb (equals black feathers) and Bw (equals white feathers).
In this kind of a cross, the heterozygous phenotype is indeed a blending of both parental types. It is easy to see how such observations would encourage the notion of the blending concept of inheritance. However, in the cross of black and white chickens or red and white flowers, only the hybrid is a phenotypic blend; the homozygous strains breed true to the parental phenotypes.
Mendel’s First Law
Mendel’s law of segregation states that in the formation of gametes, paired factors specifying alternative phenotypes (visible traits) segregate independently of one another. In one of Mendel’s original experiments, he pollinated pure-line tall plants with the pollen of pure-line dwarf plants. Thus the visible characteristics, or phenotypes, of the parents were tall and dwarf. Mendel found that all progeny in the first generation (F1) were tall, just as tall as the tall parents of the cross. The reciprocal cross—dwarf plants pollinated with tall plants—gave the same result. The tall phenotype was observed in progeny no matter which way the cross was made. Obviously, this kind of inheritance was not a blending of two traits, since none of the progeny was of intermediate size.
Next Mendel self-fertilized (“selfed”) the tall F1 plants and raised several hundred progeny, the second (F2) generation. This time, both tall and dwarf plants appeared. Again, there was no blending (no plants of intermediate size), but the appearance of dwarf plants from all tall parental plants was surprising. The dwarf trait, present in the grandparents but not in the parents, had reappeared. When he counted the actual number of tall and dwarf plants in the F2 generation, he discovered that there were almost exactly three times more tall plants than dwarf ones.
Mendel then repeated this experiment for the six other contrasting traits that he had chosen, and in every case he obtained ratios very close to 3:1 (see Figure 5-1). At this point it must have been clear to Mendel that he was dealing with hereditary determinants for the contrasting traits that did not blend when brought together. Even though the dwarf trait disappeared in the F1 generation, it reappeared fully expressed in the F2 generation. He realized that the F1 generation plants carried determinants (which he called “factors”) of both tall and dwarf parents, even though only the tall trait was expressed in the F1 generation.
Mendel called the tall factor dominant and the short recessive. Similarly, the other pairs of traits that he studied showed dominance and recessiveness. Whenever a dominant factor is present, the recessive one cannot produce its effect. The recessive trait will appear only when both factors are recessive, or in other words, in a pure condition.
In representing his crosses, Mendel used letters as symbols; dominant traits were represented by capital letters, and for recessive traits he used the corresponding lowercase letters. Modern geneticists still follow this custom. Thus the factors for pure tall plants might be represented by t/t, the pure recessive by t/t, and the mix, or hybrid, of the two plants by t/t. The slash mark is to indicate that the alleles are on homologous chromosomes. The zygote bears the complete genetic constitution of the organism. All the gametes produced by t/t must necessarily be T, whereas those produced by t/t must be t. Therefore a zygote produced by union of the two must be t/t, or a heterozygote. On the other hand, the pure tall plants (t/t) and pure dwarf plants (t/t) are homozygotes, meaning that the paired factors (alleles) are alike on the homologous chromosomes. A cross involving only one pair of contrasting traits is called a monohybrid cross.
In the cross between tall and dwarf plants there were two phenotypes: tall and dwarf. On the basis of genetic formulas there are three hereditary types: T/T, T/t, and t/t. These are called genotypes. A genotype is an allelic combination (T/T, T/t, or t/t), and the phenotype is the corresponding appearance of the organism (tall or dwarf).
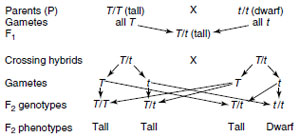 |
One of Mendel’s original crosses (tall plant and dwarf plant) could be represented as follows:
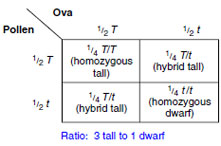 |
The next step was an important one because it enabled Mendel to test his hypothesis that every plant contained nonblending factors from both parents. He self-fertilized the plants in the F2 generation; that is, the stigma of a flower was fertilized by the pollen of the same flower.
 |
This experiment showed that the dwarf plants were pure because they at all times gave rise to short plants when self-pollinated; the tall plants contained both pure tall and hybrid tall. It also demonstrated that, although the dwarf trait disappeared in the F1 plants, which were all tall, dwarfness appeared in the F2 plants.
Mendel reasoned that the factors for tallness and dwarfness were units that did not blend when they were together. The F1 generation contained both of these units or factors, but when these plants formed their germ cells, the factors separated so that each germ cell had only one factor. In a pure plant both factors were alike; in a hybrid they were different. He concluded that individual germ cells were always pure with respect to a pair of contrasting factors, even though the germ cells were formed from hybrid individuals possessing both contrasting factors.
This idea formed the basis for his law of segregation, that is, that whenever two factors are brought together in a hybrid, they segregate into separate gametes that are produced by the hybrid. Either one of the paired factors or alleles of the parent pass with equal frequency to the gametes. We now understand that the factors segregate because there are two alleles for the character, one on each chromosome of a homologous pair, but the gametes receive only one of each in meiosis. Thus in current usage the law of segregation refers to the parting of homologous chromosomes during meiosis.
Mendel’s great contribution was his quantitative approach to inheritance. This really marks the birth of genetics, because before Mendel, people believed that traits were blended like mixing together two colors of paint, a notion that unfortunately still lingers in the minds of many and was a problem for Darwin’s theory of natural selection when he first proposed it. If traits were blended, variability would be lost in hybridization between individuals. With particulate inheritance, on the other hand, different variations are retained and can be shuffled about and resorted like blocks.
In not reporting conflicting findings, which must surely have arisen as they do in any original research, Mendel has been accused of “cooking” his results.The chances are, however, that he carefully avoided ambiguous material to strengthen his central message, which we still regard as an exemplary achievement in experimental analysis.
Testcross
When one of the alleles is dominant, heterozygous individuals are identical in phenotype to individuals homozygous for the dominant allele. Therefore you cannot determine the genotypes of these individuals just by observing their phenotypes. For instance, in Mendel’s experiment of tall and dwarf traits, it is impossible to determine the genetic constitution of the tall plants of the F2 generation by mere inspection of the tall plants. Three-fourths of this generation are tall, but which of them are heterozygotes?
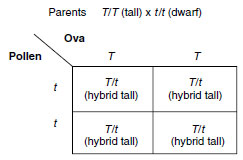 |
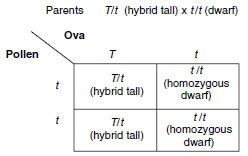 |
The testcross is often used in modern genetics for the analysis of the genetic constitution of the offspring, as well as for a quick way to make desirable homozygous stocks of animals and plants.
Intermediate Inheritance
In some cases neither allele is completely dominant over the other, and the heterozygous phenotype appears either intermediate between or even quite distinct from those of the parents. This is called intermediate inheritance, or incomplete dominance. In the four-o’clock flower (Mirabilis), two allelic variants determine red versus pink or white flowers; homozygotes are red or white flowered,
 |
When neither of the alleles is recessive, it is customary to represent both by capital letters and to distinguish them by the addition of a “prime”sign (B') or by superscript letters, for example,Bb (equals black feathers) and Bw (equals white feathers).
In this kind of a cross, the heterozygous phenotype is indeed a blending of both parental types. It is easy to see how such observations would encourage the notion of the blending concept of inheritance. However, in the cross of black and white chickens or red and white flowers, only the hybrid is a phenotypic blend; the homozygous strains breed true to the parental phenotypes.
Support our developers




