The Teleostei
The osseous fishes are occasionally devoid of any exoskeleton. Sometimes they present scattered dermal plates of true bone; or, as in the Trunkfishes (Ostracion), the body may be encased in a complete cuirass, which is calcified, but has not the structure of bone. Again, as in the Filefishes (Balistes), the skin may be beset with innumerable small spines, somewhat like those which form the shagreen of the Elasmobranchs in appearance, though they differ from them in structure. But, usually, the exoskeleton of the Teleosteans takes the form of overlapping scales, which rarely exhibit the lacunae characteristic of true bone. The free portions of the scales are sometimes smooth, and rounded at the edge, when they are termed cycloid; or they are roughened with ridges and minute spines, when they are called ctenoid.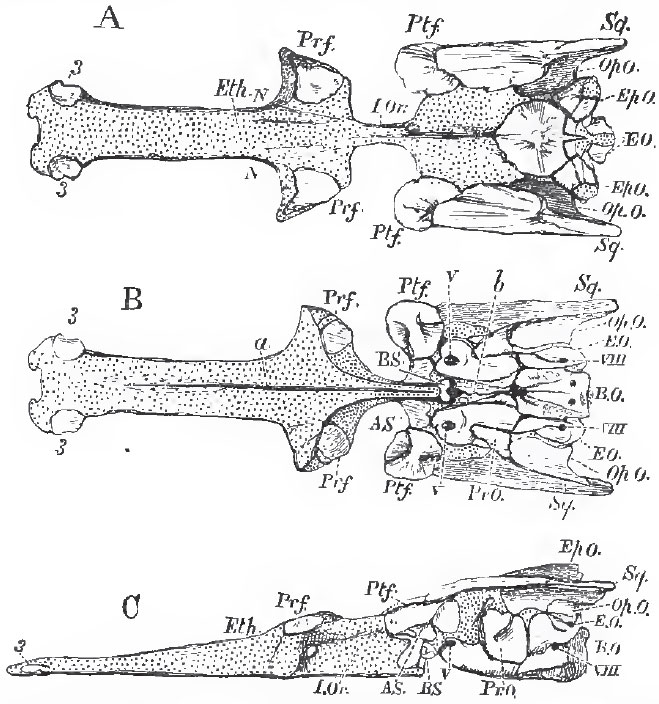 |
| Fig. 44. - The cartilaginous cranium of the Pike (Esox lucius), with its intrinsic ossifications; viewed, A, from above; B, from below; C, from the left side: N, N, nasar fossae; I. Or, Interorbital septum; a, groove for the median ridge of the parasphenoid; b, canal for the orbital muscles. Sq., wrongly so marked, is the Pterotic. V. and VIII. mark the exits of the fifth and pneumogastric nerves; 3, 3, small ossifications of the rostrum. |
The vertebrae are distinguishable only into those of the trunk and those of the tail. The latter are provided with complete inferior arches traversed by the caudal artery and vein. The former usually possess ribs, but these do not unite with one another, nor with any sternum, in the ventral median line, and they enclose the thoracico-abdominal viscera. The vertebrae are commonly united by zygapophyses, or oblique processes, placed above the centra; in addition to which, the lower margins of the centra are, not unfrequently, united by additional articular processes. Transverse processes commonly exist, but the ribs are articulated with the bodies of the vertebrae, or with the bases of the transverse processes, not with their extremities.
When a dorsal fin exists in the trunk, its rays are articulated with, and supported by, elongated and pointed bones - the interspinous bones, which are developed around preexisting cartilages, and lie between, and are connected with, the spines of the vertebrae. The fin-rays may be entire and completely ossified, or they may be transversely jointed and longitudinally subdivided at their extremities. Not unfrequently, the articulation between the fin-rays and the interspinous bone is effected by the interlocking of two rings-one belonging to the base of the fin-ray and its included dermal cartilage, and the other to the summit of the interspinous bone-like the adjacent links of a chain.
In all Teleostean fishes the extremity of the spinal column bends up, and a far greater number of the caudal fin-rays lie below than above it. These fishes are, therefore, strictly speaking, heterocercal. Nevertheless, in the great majority of them (as has been already mentioned, page 19), the tail seems, upon a superficial view, to be symmetrical, the spinal column appearing to terminate in the centre of a wedge-shaped hypural bone, to the free edges of which the caudal fin-rays are attached, so as to form an upper and a lower lobe, which are equal, or subequal. This characteristically Teleostean structure of the tail-fin has been termed homocercal-a name which may be retained, though it originated in a misconception of the relation of this structure to the heterocercal condition.
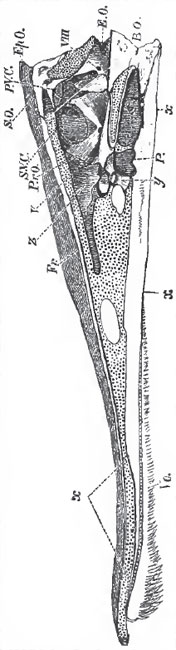 |
| Fig. 45. - Longitudinal and vertical section of a fresh Pike'a skull. - The cut surface of ourttilage is dotted. For S. V. C. and P.V.G., read a.s.c., anterior, and p.s.c., posterior semicircular canal; x, the parasphenoid; y, the basi sphenoid; Vo, the vomer P., the pituitary fossa. |
In no Teleostean fish is the bent-up termination of the notochord replaced by vertebrae. Sometimes, as in the Salmon (Fig. 6, page 20), it becomes ensheathed in cartilage, and persists throughout life. But, more usually, its sheath becomes calcified, and the urostyle thus formed coalesces with the dorsal edge of the upper part of the wedge-shaped hypural bone, formed by the anchylosis of a series of ossicles, which are developed in connection with the ventral face of the sheath of the notochord.
In the caudal region of the body, interspinous bones are developed between the spines of the inferior arches of the vertebrae, and bear the fin-rays of the anal, and, in part, of the caudal fin.
The Teleostei differ very much in the extent to which the primordial cranium persists throughout life. Sometimes, as in the Pike (Figs. 44 and 45), it grows with the growth of the fish, and only becomes partially ossified; in other cases it almost disappears. A basi-occipital (B. 0.), ex-ocoipital (E. 0.), and supra-occipital (S. 0.) bone are developed in it, and form a complete occipital segment. The proper basi-sphenoid (BS) bone is always a very small, and usually somewhat Y-shaped, bone. The alisphenoids (AS.) sometimes are and sometimes are not developed. The presphenoidal and orbitosphenoidal regions commonly, but not always, remain unossified.
In most osseous fishes, the base of the skull in front of the basisphenoid is greatly compressed from side to side, and forms an interorbital septum (I Or.). The anterior moiety of the cranial cavity is consequently reduced to a comparatively narrow passage above the septum (Fig. 45). In the Siluroid and Cyprinoid fishes, however, this septum is not formed, and the cranial cavity is of nearly equal size throughout, or gradually diminishing forward. The ethmoidal cartilage usually remains unossified, but sometimes, as in the Pike, ossification may take place in it. (Fig. 44, 3, 3.) The antorbital, or lateral ethmoidal, processes of the primordial cranium ossify, and give rise to the prefrontal bones (Prf.). The postorbital processes also ossify as postfrontals (Ptf.). The upper and posterior part of the primordial cranium exhibits five processes-one postero-median, two postero- lateral, and two postero-external. The postero-median ossifies as part of the supra-occipital (S. 0.). The postero-lateral ossifies as part of the epiotic (Ep. 0.), which lies upon the summit of the superior vertical semicircular canal. The postero-external closely corresponds with the squamosal of the higher Vertebrata in position; but, as a cartilage bone, it corresponds with an ossification of the capsule of the ear, called pterotic in the higher Vertebrata. Not unfrequently, as in the Cod, for example, the opisthotio (Op. O.) is a distinct bone, and enters into the formation of the postero-external process. The prootio (Pr. 0.) is always a welldeveloped bone, and occupies its regular place, in front of the anterior vertical semicircular canal, and behind the exit of the trigeminal nerve.
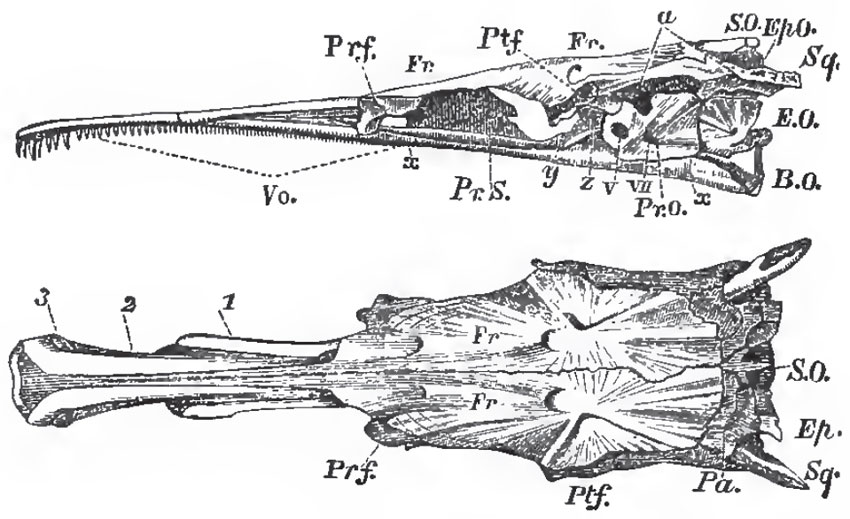 |
| Fig. 46. - Side and upper views of the skull of a Pike (Esox lucius), without the facial or supra-orbital bones: y, the basisphenoid; z, the alisphenoid; a, the articular fecet for the hyomandibular bone. |
1. The parietal bones (Pa.), which sometimes meet in a sagittal suture, as in most of the higher Vertebrata, but are very generally separated by the junction of the frontals with the supra-occipital.
2. The large frontals (Fr.), which may or may not unite into one.
3. The nasal bones (Na.), apparently replaced in the Pike by the bones 1 and 2.
The under-surface of the skull possesses two membrane bones: in front the vomer (Vo.), and, behind, the huge parasphenoid (x, x), which ensheathes all the basis cranii, from the basi-occipital to the vomer.
A supra-orbital bone (S. Or.) is the only membrane bone attached to the sides of the brain-case. Two premaxillary bones (Pmx.) are attached, sometimes closely, sometimes loosely, to the anterior extremity of the cranium; and behind these are the maxillae (Mx.), which are sometimes large and single, as in the Cyprinoid fishes, but may become subdivided, or be reduced to mere styliform supports for cirri, as in many Siluroid fishes. In most osseous fishes the maxillae take little or no share in the formation of the gape, which is bounded above by the backwardly-extended premaxillae.
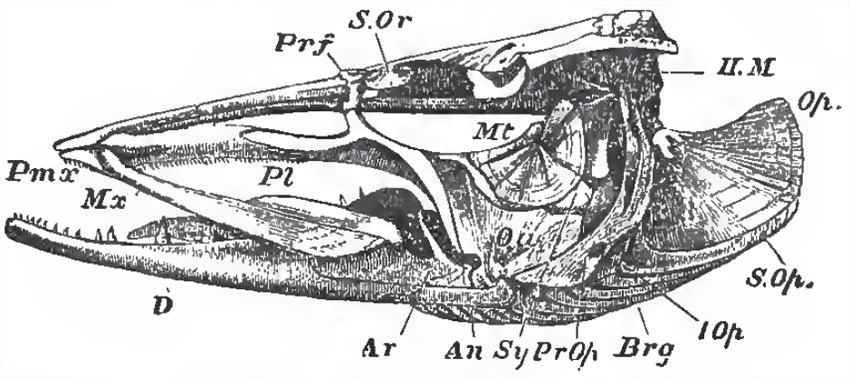 |
| Fig. 47. - Side-view of the skull of a Pike (Esox lucius): Prf, prefrontal; II. M hyomandibular bone; Op, operculum; S.Op., suboperculum; L. Op, interoperculum; Pr, Op, preoperculum; Brg, branchiostegal rays; Sy, symplectic; Mt, metapterygoid; Pl, palato-pterygoid arch; Qu, quadrate bone; Ar, articular; An, angular; D, dentary: S.Or, suborbital bone. |
The palato-quadrate and hyomandibular have essentially the same structure and arrangement as in Lepidosteus and Amia, The homologue of the suspensorium of the Elasmobranchii is articulated with a surface furnished to it by the postfrontal, pterotic, and prootic bones. Usually it moves freely upon that surface, but, in the Plectognathi, it may be fixed. It ossifies so as to give rise to two bones: an upper broad hyomandibular (H.M.), with which the operculum articulates; and a lower styliform symplectic (Sy.), which fits into a groove on the inner and posterior surafce of the quadrate, and is firmly held there.
The palato-quadrate arch is represented by several bones, of which the most constant are the palatine (Pl.) in front, and the quadrate (Qu.) behind and below. Besides these there may be three others: an external, ectopterygoid (Ecpt.), an internal, entopterygoid (Ept.), and a metapterygoid (Mpt.). The last envelops the upper and posterior portion of the primitive quadrate cartilage; and, fixing itself against the hyomandibular, contributes to the firmness of the union already effected by the symplectic.
Meckel's cartilage (Mck.) persists throughout life, but the ossification of its proximal end gives rise to an os articulare in the lower jaw. To these an angular (An.) and a dentary (D.) membrane bone are commonly added (Fig. 47).
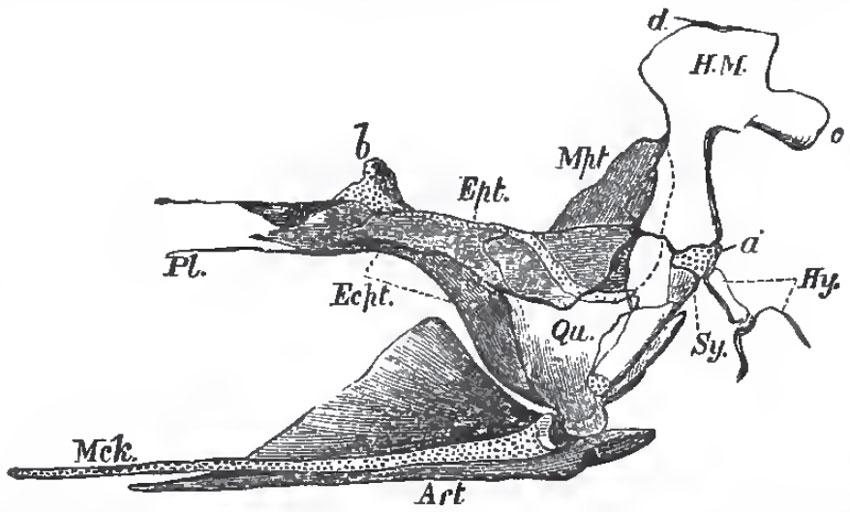 |
| Fig. 48. - Palato-quadrate arch, with the hyomandibular and symplectic of the Pike, viewed from the inner side; the articular piece (Art), of the lower jaw, and Meckel's cartilage (Mck.) of the Pike; seen from the inner side: a, the cartilage interposed between the hyomandibular (H.M.), and the symplectic (Sy); b, that which serves as a pedicle to the pterygo-palatine arch; c, process of the hyomandibular with which the operculum articulates; d, head of the hyomandibular which articulates with the skull. |
The hyoidean arch is usually composed of two large cornua - connected with the cartilaginous interval between the hyomandibular and the symplectic by a stylohyal ossification, and abutting, in the middle line below, upon one or more median pieces, the anterior of which (entoglossal) supports the tongue, while the posterior (urohyal) extends back to join the median elements of the branchial apparatus. The cornua themselves are usually ossified into four pieces: an upper (epihyal) and a lower (ceratohyal) large ossification, and two small ones (basihyals) connected with the ventral ends of the lower large ossification.
There are usually five pair of branchial arches connected by median ventral ossifications. The posterior pair are single bones, which underlie the floor of the pharynx, bear no branchial filaments, but commonly support teeth, and are called hypopharyngeal bones. In certain osseous fishes, thence called Pharyngognathi, they anchylose together into one bone. The anterior four pair are composed of several joints, and the uppermost articulations of more or fewer of them usually expand, bear teeth, and form the epipharyngeal bones. Sundry important membrane bones are connected with the mandibular and hyoidean arches. The preoperculum, (P. Op.), operculum (Op.), and branchiostegal rays, (Br.), already met with among the Ganoidei, are the most constant of these. Beneath the operculum, lies a suboperculum (S. Op.), and below this an interoperculum (I. Op.), which is connected by ligament with the angular piece of the lower jaw, and is also united to the outer face of the hyoidean arch. It may be altogether ligamentous, as in the Siluroids.
The branchiostegal rays are attached partly to the inner, and partly to the outer, surface of the hyoidean arch. They support a membrane, the branchiostegal membrane, which serves as a sort of inner gill-cover.
Most Teleostei possess two pair of limbs, the pectoral and the ventral fins. But the latter are often absent, and the former are occasionally wanting. When the pectoral fins are absent, the pectoral arch usually remains, though it may be reduced to little more than a filament, as in Muraenophis, The ventral fins are frequently situated in their normal position beneath the posterior part of the trunk; but in considerable groups of these fishes they are immediately behind the pectoral fins (thoracic), or even in front of them (jugular). In the asymmetrical Pleuronectidae one pectoral fin may be larger than the other, or may alone remain, as in Monochirus.
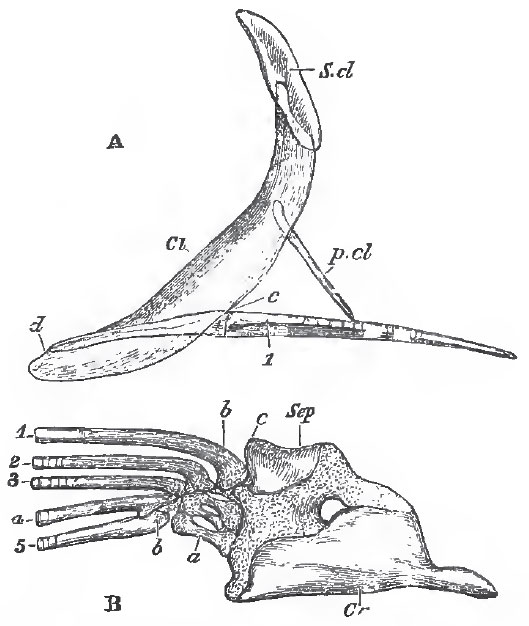 |
| Fig. 49. - The bones of the pectoral arch and fore-limb of the Pike (Esox lucius): A, a
semi-diagrammatic view of these bones, to show their relative natural position. The
clavicle (Cl) is supposed to be transparent. S.cl, supra-clavicula; p.cl, post-clavicula c, d, the posterior and anterior ends of the outer margin of the scapulo-coracoid. - B, the
scapulo-coracoid and limb separate and on a larger scale; Scp, scapula; Or, coracoid a, basal cartilages; b, fin-rays; c, corresponds with c in the foregoing figure. |
The pectoral arch always consists of a primarily cartilaginous coraco-scapular portion - which usually ossifies in two pieces, a coracoid below, and a scapula above - and of sundry membrane bones. The chief of these membrane bones is the clavicula (Cl), which meets its fellow in the middle line, and is usually joined to it by ligament, but sometimes, as in the Siluroids, by sutural union. By its inner surface it gives attachment to the coraco-scapular - and sometimes above them, to a styliform bone which extends back among the lateral muscles-the post-clavicula (p.cl.).
Attached to the dorsal end of the clavicle, there is usually a second much smaller bone, the supra-clavicula (S.cl.), and this is very generally connected with the skull by a superficial membrane bone, the post-temporal, which, in front, becomes forked, and attaches itself by one prong to the epiotic bone, by the other to the pterotio, or lower down to the side of the cranium. The base of the fin contains a series of not more than five, more or less ossified, cartilages, which are placed side by side and articulate with the coraco-scapular; to these succeed one or more rows of small cartilages, partially hidden by the bases of the exoskeletal fin-rays. The most anterior of these basal cartilages (the mesopterygial basale) is enclosed by the base of the anterior fin-ray, and effects that articulation with the shoulder-girdle which is so remarkable in many solenoid fishes. The posterior cartilage, or bone, is the metapterygial basale, and the intermediate three are radialia (p. 39).
Most Teleostei possess teeth, and, in the majority of these fishes, teeth are very widely distributed over the surface of the walls of the oral and pharyngeal cavities. The teeth very very much in structure; ordinarily, they consist of dentine, capped with structureless enamel. The parietes of the tooth are not unfrequently longitudinally folded toward the base, but this folding never goes so far as in the Ganoids. The different kinds and modes of arrangement of the teeth may be classified as follows:
1. Isolated, more or less pointed teeth, developed from papillae of the mucous membrane, which do not become enclosed in sacs-frequently anchylosed to the subjacent bone, but not imbedded in alveoli, nor replaced vertically. The great majority of ordinary osseous fishes have teeth of this kind.
2. Isolated teeth, which become imbedded in sockets, and are replaced vertically.
Such teeth are seen in the premaxillae of Sargus, where they curiously simulate the form of human incisors; and, imbedded in the coalesced hypopharyngeal bones, in Labrus.
3. Isolated teeth, imbedded in the substance of the bone which supports them. The teeth and the supporting bone wear away in front, and are replaced by new teeth developed behind the others. This structure is seen in the coalesced hypopharyngeal bones of the Parrotfish (Scarus).
4. Beak-like compound teeth, attached to the premaxillae and dentary bones of the mandible.
These are of two kinds. In the Parrotfish (Scarus) the beak is formed by the union of numerous separately-developed teeth into one mass. But in the Gymnodonts (Tetrodon and Diodon) the beak is produced by the coalescence of broad calcified horizontal lamellae thrown off from a subjacent pulp.
5. In the Carp and its allies the basi-occipital sends down a median process, which expands at the end, and supports a broad, thick, horny tooth.
The stomach is usually wide and sac-like, but sometimes (in Scomberesoces, Cyprinoids, and others) is not wider than the intestine. Occasionally, as in Mugil, it acquires thick walls and becomes gizzard-like. The commencement of the small intestine is very generally marked by the presence of more or less numerous caecal diverticula, the pyloric caeca. The small intestine has no spiral valve, though the mucous membrane may be raised into large transverse folds. The rectum does not terminate in a cloaca, and almost always opens quite separately from the urinary and genital ducts, and in front of them.
In many Teleostean fishes an air-bladder underlies the vertebral column, and is connected by an open pneumatic duct with the dorsal wall of the oesophagus, or even with the stomach, as in the Herring. In other Teleostei, the air-bladder occupies the same position, but is closed, the duct by which the air-bladder is primitively connected with the alimentary canal becoming obliterated. In a comparatively small number of the Teleostei-the Blennii, the Pleuronectidae or Flatfishes, the Sand-eel (Ammodytes), the Loricarini, and Symbranchii, and some members of other families-there is no air-bladder. In those Teleostei in which it is present, it may be divided into two parts by a constriction; or it may be prolonged into diverticula; or retia mirabilia may be developed in its walls. Sometimes the air-bladder is brought into direct relation with the membranous labyrinth, as in Myripristis and Sparus, and the Herring, Shad, and Anchovy-prolongations of the one organ being separated from the other only by a membranous fenestra in the wall of the skull. In the Siluroidei, Cyprinoidei, and Characini, and in the Gymnotini, the anterior end of the air-bladder is connected with the membranous vestibule by the intermediation of a series of bones attached to the vertebral column, some of which are movable.
The vessels of the air-bladder are derived from, and empty themselves into, those of the adjacent parts of the body, in which respect, and in the dorsal position of the oesophageal aperture of the pneumatic duct, this structure differs from a lung.
The heart consists of a single auricle, receiving its blood from a venous sinus; and of a single ventricle, separated by a single row of valves from the bulbus aortoe, which is not rhythmically contractile.
The cardiac aorta divides into trunks to form the branchial arteries, which run upon the outer, or convex, side of the branchial arches, and are distributed to the branchial filaments. The blood is collected thence into a branchial vein, which also lies on the convex side of the arch; and, increasing toward its dorsal end, opens into one of the trunks of the original dorsal aorta. Of these there are two, a right and a left, which pass backward and meet in the trunk of the dorsal aorta under the spinal column.
The anterior branchial vein gives off, at its dorsal termination, a considerable carotid trunk, which passes forward under the base of the skull; and this is united with its fellow by a transverse branch-so that a complete arterial circle, the circulus cephalicus, is formed beneath the base of the skull. Below, the anterior branchial vein gives off the hyoidean artery, which ascends along the hyoidean arch, and very generally terminates by one branch in the cephalic circle, and by another enters a rete mirabile, which lies in the inner side of the hyomandibular bone, and sometimes has the form of a gill. This is the pseudobranchia. The branches of the rete mirabile unite again into the ophthalmic artery, which pierces the sclerotic, and breaks up into another rete mirabile, the choroid gland, before being finally distributed.
In the Lamprey, as has been seen, the respiratory organs are pouches, the anterior and posterior walls of which are raised into vascular folds. The walls of adjacent pouches are distinct and but loosely connected together; and considerable spaces of integument separate their rounded outer apertures.
In the ordinary Elasmobranchii, the branchial pouches are more flattened from before backward, and their outer apertures are more slit-like. The integumentary spaces between the slits are correspondingly narrower, and the adjacent walls of successive pouches are more closely approximated, so that they are divided only by sepia; but the vascular plaits of the surface of the respiratory mucous membrane do not reach the outer edges of these septa.
In Chimaera, the free edges of the septa are exceedingly narrow, and the apices of the branchial processes extend outward to them.
In the Sturgeon, the septum is not more than three-fourths as long as the branchial processes, the apices of which are consequently free.
The process of reduction is carried still further in the Teleostei-the septum not attaining to more than one-third the length of the branchial processes; and, as in the Ganoids, each process is supported by an osseous or cartilaginous skeleton.
The Teleostei have no functional hyoidean, or opercular, gill; and, as a general rule, each of their four branchial arches possesses a double series of branchial processes, making eight in all. Not unfrequently (Cottus, Cyclopterus, Zeus, etc.), the number is reduced to seven; the fourth branchial arch having only one series, the anterior. In this case, the gill-cleft, which should lie between this arch and the fifth, is closed. Sometimes there are only six series of branchial processes, the fourth arch being devoid of any (e. g., Lophius, Diodon). In Malthaea the number is reduced to five, only the anterior series of the third arch being developed; and in Amphipnous cuchia only the second branchial arch possesses branchial filaments, the first, third, and fourth, being devoid of them.
Many Teleostean fishes possess accessory respiratory organs. These may take the form of arborescent appendages to the upper ends of some of the branchial arches, as in Clarias, Heterobranchus, and Heterotis; or, as in the Climbing Perch (Anabas) and its allies, the epipharyngeal bones may enlarge and acquire a labyrinthic honeycombed structure, and support a large surface of vascular mucous membrane; or, as in the Clupeoid (Lutodeira chanos), an accessory gill may be developed in a curved caecal prolongation of the branchial cavity. Finally, in Saccobranchus singio and in Amphipnous cuchia, the membrane lining the branchial chamber is prolonged into sacs, which lie at the sides of the body, and receive the blood from the divisions of the cardiac aorta which supply the branchise, while they return it into the dorsal aorta.
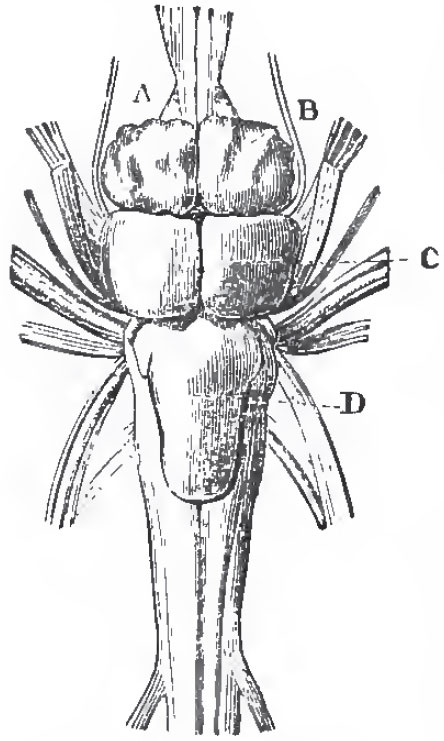 |
| Fig. 50. - Brain of tlie Pike, viewed from above: A, the olfactory nerves or lobes, and beneath them the optic nerves; B, the cerebral hemispheres; C, the optic lobes; D, the cerebellum. |
All these fishes (except Lutodeira) are remarkable for their power of sustaining life out of the water. Many inhabit the marshes of hot countries, which become more or less desiccated in the dry season.
The kidneys of Teleostean fishes receive a great part of their blood from the caudal vein, which ramifies in them. They vary greatly in length, sometimes extending along the whole under-surface of the vertebral column, from the head to the termination of the abdomen. The ureters pass into a urinary bladder which opens behind the rectum.
The brain in the Teleostei has solid cerebral hemispheres, and, when viewed from above, the thalamencophalon is hidden by the approximation to the hemispheres of the large and hollow optic lobes of the mesencephalon, which has a pair of inferior enlargements, lobi inferiores. There is a peculiarity about the structure. of the optic lobes, which has given rise to much diversity of interpretation of the parts of the brain in osseous fishes. The posterior wall of these lobes, where it passes into the cerebellum, or in the region which nearly answers to the valve of Vieussens in mammals, is thrown forward into a deep fold which lies above the crura cerebri, and divides the iter a tertio ad quartum ventriculum from the ventricle of the optic lobes throughout almost the whole extent of the latter. This fold is the "fornix" of Gottsche. On each side of it the floor of the ventricle of the optic lobes is raised up into one or more eminences, which have the same relation to the optic lobes as the corpora striata have to the prosencephalic vesicle.
The optic nerves simply cross one another, and form no chiasma. The cerebellum is usually large.
The cephalic part of the sympathetic nerve is present, as in the higher Vertebrata.
Each of the nasal sacs usually opens externally by two apertures. In some Gymnodonts a solid tentacle is said to take the place of a nasal sac.
The eyes are abortive in the Blind-fish of the caves of Kentucky (Amblyopsis spelceus). A fibrous band often passes from the back of the orbit to the sclerotic, and represents the cartilaginous pedicle of the Elasmobranchs. There is no nictitating membrane, but immovable external eyelids may be developed. The choroidal gland, mentioned above, surrounds the optic nerve between the sclerotic and the choroid. Very generally, a falciform process of the latter membrane traverses the retina and vitreous humor to the crystalline lens. This represents the pecten of higher Vertebrata. As in other fishes, the lens is spheroidal, and the cornea flat. The sacculus of the auditory organ contains large solid otoliths, which are usually two in number - the larger, anterior one, is termed Sagitta; the smaller, posterior, Asteriscus. There are always three large semicircular canals.
The reproductive organs are either solid glands which burst into the abdominal cavity, whence their reproductive elements are conveyed away by abdominal pores; or, as is more usual, they are hollow organs, and are continued backward into ducts which open beside, or behind, the urinary aperture.
Some few Teleostei are ovoviviparous (e. g., Zoarces viviparus), the eggs being retained in the interior of the ovary, and hatched there. In the male Syngnathus, and other Lophohranchii, integumentary folds of the abdomen grow down and form a pouch, into which the eggs are received, and in which they remain until they are hatched.
The young of osseous fishes are not known to undergo any metamorphosis, nor are they provided with external gills, nor with spiracula.
The classification of the Teleostei is not yet in a thoroughly satisfactory state, and the following arrangement must be regarded as provisional:
1. The Physostomi.- This group contains the Siluroidei, the Cyprinoidei, the Characini, the Cyprinodontes, the Salmonidoe, the Scopelini, the Esocini, the Mormyri, the Galaxice, the Clupeidae, the Heteropygii, the Muraenoidei, Symbranchii, and Gymnotini. The air-bladder is almost always present, and, when it exists, has an open pneumatic duct. The skin is either naked, or provided with bony plates, or cycloid scales; the ventral fins, when present, are abdominal in position. The fin-rays (except in the pectoral and dorsal fins of sundry Siluroidei) are all soft and jointed. The inferior pharyngeal bones are always distinct.
In all other Teleostean fishes the air-bladder is either absent, or devoid of an open pneumatic duct. Hence they are termed, collectively, Physoclisti by Haeckel.
2. The Anacanthini. - The body has cycloid or ctenoid scales, or is naked. The ventral fins, if present, are jugular in position. The fin-rays are all articulated. The inferior pharyngeal bones are distinct. (Ophidini, Gadoidei, Pleuronectidae.)
The Pleuronectidae are the most aberrant of all Teleostean fishes, on account of the disturbance in the bilateral symmetry of the body, skull, and fins, to which reference has already been made (p. 30).
3. The Acanthopteri have generally ctenoid scales, thoracic or jugular ventral fins, entire fin-rays in some of the fins, and distmct inferior pharyngeal bones. The Percoidei, Cataphracti, Sparoidei, Scicenoidei, Labyrinthici, Mugiloidei, Notacanthini, Scomberoidei, Squamipennes, Taenioidei, Gobioidei, Blennioidei, Pediculati, Theuthyes, and Fistulares, belong to this great group.
4. The Pharyngognathi is the name given by Muller to a somewhat artificial assemblage of fishes, the only common characters of which are the anchylosis of the inferior pharyngeal bones and the closed pneumatic duct. They have either cycloid or ctenoid scales. The ventral fins may be abdominal or thoracic. The anterior dorsal and ventral fin-rays may be either unjointed, as in the Labroidei, Pomacentridae, Chromidae; or articulated, as in the Scomberesoces.
The two remaining groups are very peculiar; but I confess. I do not see upon what ground they can be regarded as of ordinal value.
5. The Lophobranchii. - The body is covered with bony plates. The ventral fins are almost always absent. The inferior pharyngeal bones are distinct. The branchial processes have a clavate form, being larger at the free than at the attached ends, and are in this respect unlike those of any other fishes. (Pegasidae, Syngnathidae.)
6. The Plectognathi. - The body is covered with plates or spines. The ventral fins are absent or represented only by spines. The inferior pharyngeal bones are distinct. The premaxillae and, usually, the hyomandibular, are immovably united with the skull-a character of rare occurrence among other fishes. (Gymnodontidae, Ostraciontidae, Balistidae.)
The greater number of Teleostei are marine. No Anacanthini, Plectognathi, or Lophobranchii, and only one family of Pharyngognathi (the Chromidae), inhabit completely fresh water. Comparatively few Acanthopteri are fluviatile. On the other hand, by far the greater number of the Physostomi are, either temporarily or permanently, fresh-water fish.
If the Leptolepidae (Thrissops, Leptolepis, Tharsis) are Ganoids, the Teleostei are not known before the Cretaceous epoch, when both Physostomi and Acanthopteri make their appearance, under forms, some of which (e. g., Beryx) are generically identical with fish living at the present day.
Support our developers




