Endosymbiosis and Origin of Eukaryotic Algae
Within the algae, different evolutionary lineages are discernable. Three major eukaryotic photosynthetic
groups have descended from a common prokaryotic ancestor, through an endosymbiotic
event. Therefore, these algae possess primary plastid, that is, derived directly from the prokaryotic
ancestor. Other algal groups have acquired their plastids via secondary (or tertiary) endosymbiosis,
where a eukaryote already equipped with plastids is preyed upon by a second eukaryotic cell. Endosymbiotic process produced nested cellular compartments one inside the other, which can give
information about the evolutionary history of the algae containing them.
Cyanobacteria evolved more than 2.8 billion years ago and have played fundamental roles in
driving much of the ocean carbon, oxygen, and nitrogen fluxes from that time to present. The evolution
of cyanobacteria was a major turning point in biogeochemistry of Earth. Prior to the appearance of these organisms, all photosynthetic organisms were anaerobic bacteria that used light
to couple the reduction of carbon dioxide to the oxidation of low free energy molecules, such as H2S
or preformed organics.
Cyanobacteria developed a metabolic process, the photosynthesis, which
exploits the energy of visible light to oxidize water and simultaneously reduces CO2 to organic
carbon represented by (CH2O)n using light energy as a substrate and chlorophyll a as a requisite
catalytic agent. Formally oxygenic photosynthesis can be summarized as:
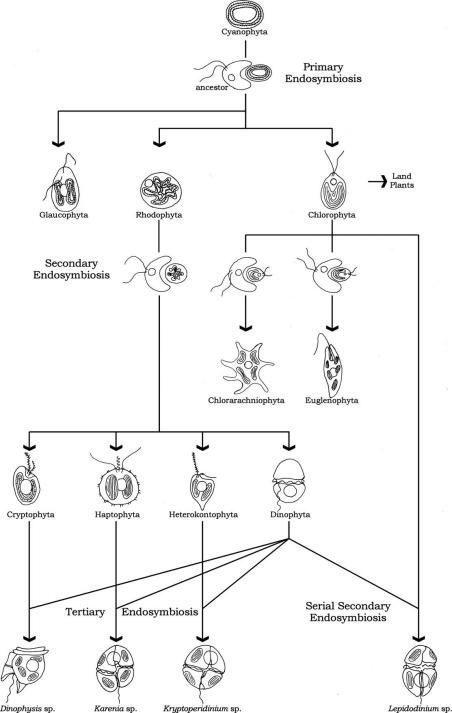
FIGURE 1.48 Algal evolution and endosymbiotic events.
All other oxygen producing algae are eukaryotic, that is, they contain internal organelles,
including a nucleus, one or more chloroplasts, one or more mitochondria, and, most importantly,
in many cases they contain a membrane-bound storage compartment or vacuole. The three
major algal lineages of primary plastids are the Glaucophyta lineage, the Chlorophyta lineage,
and the Rhodopyta lineage (Figure 1.48).
FIGURE 1.48 Algal evolution and endosymbiotic events.
Glaucophyta lineage occupies a key position in the evolution of plastids. Unlike other plastids,
the plastids of glaucophytes retain the remnant of a Gram-negative bacterial cell wall of the type
found in cyanobacteria,
with a thin peptidoglycan cell wall and cyanobacterium-like pigmentation
that clearly indicate its cyanobacterial ancestry. In fact, the
Cyanophora paradoxa plastid genome
shows the same reduction as other plastids when compared with free-living cyanobacteria (it is
136 kb and contains 191 genes). The peptidoglycan cell wall of the plastid is thus a feature retained
from their free-living cyanobacterial ancestor. In this context, the Glaucophyta are remarkable only
for their retention of an ancestral character present in neither green nor red plastids. No certain case
of a secondary plastid derived from Glaucophyta is known.
Green algae (Chlorophyta) constitute the second lineage of primary plastids. The simple twomembrane
system surrounding the plastid, the congruence of phylogenies based on nuclear and
organellar genes, and the antiquity of the green algae in the fossil record all indicate that the
green algal plastid is of primary origin. In these chloroplasts, chlorophyll b was synthesized as a
secondary pigment and phycobiliproteins were lost. Another hypothesis is that the photosynthetic
ancestor of green lineage was a prochlorophyte that possessed chlorophylls a and b and lacked
phycobiliproteins.
The green lineage played a major role in oceanic food webs and the carbon cycle from about 2.2
billion years ago until the end-Permian extinction, approximately 250 million years ago. It was this
similarity to the pigments of plants that led to the inference that the ancestors of land plants (i.e.,
embryophytes) would be among the green algae, and is clear that phylogenetically plants are a
group of green algae adapted to life on land. Euglenophyta and Chlorarachniophyta are derived
from this primary plastid lineage by secondary endosymbiosis; the green algal plastid present in
Euglenophyta is bounded by three membranes, while the green algal plastid present in the Chlorarachniophyta
is bound by four membranes.
Since that time, however, a second group of eukaryotes has risen to ecological prominence; that
group is commonly called the “red lineage.” The plastids of the red algae (Rhodophyta) constitute
the third primary plastid lineage. Like the green algae, the red algae are an ancient group in the
fossil record, and some of the oldest fossils interpreted as being of eukaryotic origin are often
referred to the red algae, although clearly these organisms were very different from any extant
alga. Like those of green algae, the plastids of red algae are surrounded by two membranes.
However, they are pigmented with chlorophyll a and phycobiliproteins, which are organized
into phycobilisomes. Phycobilisomes are relatively large light-harvesting pigment/protein
complexes that are water-soluble and attached to the surface of the thylakoid membrane.
Thylakoids with phycobilisomes do not form stacks like those in other plastids, and consequently
the plastids of red algae (and glaucophytes) bear an obvious ultrastructural resemblance to
cyanobacteria.
A number of algal groups have secondary plastids derived from those of red algae, including
several with distinctive pigmentation. The cryptomonads (Cryptophyta) were the first group in
which secondary plastids were recognized on the basis of their complex four membrane structure.
Like red algae, they have chlorophyll a and phycobiliproteins, but these are distributed in the
intrathylakoidal space rather than in the phycobilisomes found in red algae, Glaucophyta, and
Cyanophyta. In addition, cryptomonads possess a second type of chlorophyll, chlorophyll
c,
which is found in the remaining red lineage plastids. These groups, which include the Heterokontophyta
(including kelps, diatoms, chrysophytes, and related groups), Haptophyta (the coccolithophorids),
and probably those dinoflagellates (Dinophyta) pigmented with peridinin, have
chlorophylls
a and
c, along with a variety of carotenoids, for pigmentation. Stacked thylakoids
are found in those lineages (including the cryptomonads) that lack phycobilisomes. The derivation of chlorophyll
c containing plastids from the red algal lineage is still somewhat conjectural, but
recent analyses of both gene sequences and gene content are consistent with this conclusion.
A few groups of dinoflagellates have plastids now recognized to be derived from serial
secondary endosymbiosis (the uptake of a new primary plastid-containing endosymbiont) such
as Lepidodinium spp. or tertiary endosymbiosis (the uptake of the secondary plastid-containing
endosymbiont), such as
Dinophysis,
Karenia, and
Kryptoperidinium.
All these groups are comparatively modern organisms; indeed, the rise of dinoflagellates and
coccolithophorids approximately parallels the rise of dinosaurs, while the rise of diatoms approximates
the rise of mammals in the Cenozoic. The burial and subsequent diagenesis of organic carbon
produced primarily by members of the red lineage in shallow seas in the Jurassic period provide the
source rocks for most of the petroleum reservoirs that have been exploited for the past century by
humans.




